











Россия
Россия
В период зимовки у судаков из естественной среды обитания и выращенных в заводских условиях происходят изменения в морфофизиологическом и химическом статусе, обусловленные расходованием энергетических резервов организма и перераспределением питательных веществ в мышцах и печени. Это выражается в значительном снижении содержания полостного жира в теле диких и заводских особей (на 45 и 16 % соответственно), жирности мышц (на 26 % у диких особей) и печени (на 59 % у диких и 16 % у заводских рыб), витамина С в мышцах (на 56 и 23 % соответственно). При этом в мышцах и печени многократно повышается содержание безазотистых экстрактивных веществ – в 2,1 и 2,8 раза соответственно у диких особей и в 1,5 и 1,4 раза у судаков, выращенных в садках. В липидах мышц судаков в зимний период существенно повышается доля мононенасыщенных жирных кислот и снижается доля полиненасыщенных жирных кислот (ПНЖК), особенно эйкозапентаеновой (ЭПК), докозагексаеновой (ДГК) и арахидоновой, на фоне повышения уровня этих кислот в печени. С точки зрения качества мяса, оцененного по содержанию длинноцепочечных ПНЖК, за период зимовки происходит значительное снижение пищевой ценности выловленных и выращенных рыб, что выражается в 2-3-кратном снижении ЭПК + ДГК в мышцах. Подобные изменения являются адаптацией рыб к холодным температурным условиям в период длительной зимовки и связаны с половым созреванием.
судак, дикие и заводские рыбы, зимовка, коэффициент упитанности, мышцы, печень, полостной жир, жирные кислоты, жирнокислотный состав, питательная ценность
Введение
В условиях современного состояния мирового промысла рыбы аквакультура рассматривается в качестве альтернативного поставщика высококачественной рыбной продукции. В России, с ее огромным фондом рек и озер, более развито пресноводное рыбоводство, в котором в качестве объектов выращивания превалируют лососевые, карповые и осетровые виды. В рамках экспериментальных исследований разрабатываются технологии выращивания окуневых рыб, среди которых судак является наиболее традиционным и экономически выгодным видом. Первые сведения о разведении судака в России относятся к 1880-м гг. и связаны с деятельностью Никольского рыбоводного завода [1]. В работе не указана технология культивирования, однако следует полагать, что она была основана на прудовом разведении судака с последующим зарыблением озер. Подобный метод разведения судака использовали в нашей стране до недавнего времени, пока с развитием современных форм индустриального рыбоводства судака не начали выращивать садковым или бассейновым способом, в том числе в установках замкнутого водоснабжения (УЗВ) [2, 3].
Введение судака в индустриальную аквакультуру имеет сложности, не решенные по настоящее время и связанные с отсутствием на рынке специализированных кормов (форелевые или осетровые корма приводят к ожирению окуневых рыб) и нехваткой знаний о физиологических особенностях судака при содержании в неволе. Например, в УЗВ созревание судака проходит с отклонениями, что обусловлено отсутствием возможности воспроизвести в заводских условиях комплекс абиотических факторов, соответствующих природным, в том числе условиям зимовки [4, 5]. Известно, что содержание объектов выращивания в зимний период является одним из проблемных мест рыбоводного процесса, которое, в силу физиологических и химических изменений в организме, может сопровождаться массовой гибелью рыб, особенно при нерациональном летнем кормлении. Таким образом, развитие технологии культивирования судака индустриальным способом должно основываться на расширении знаний о его биологии и физиологии в различные периоды рыбоводного цикла.
Помимо биологических параметров, особый интерес представляет исследование питательной ценности рыб в зависимости от сезона выращивания, что может способствовать рациональному ведению рыбного хозяйства и получению рыбоводной продукции более высокого качества.
В связи с вышесказанным целью настоящей работы явилось исследование морфофизиологических и биохимических параметров диких и искусственно выращенных судаков до и после зимовки.
Материал и методы
Объектом исследования служили неполовозрелые судаки (четырехлетки и четырехгодовики) из оз. Суходольское и соразмерные с дикими рыбами искусственно выращенные особи (трехлетки и трехгодовики). Последние получены из ремонтно-маточного стада, содержащегося в садках рыбоводного хозяйства ООО «Форват» (Приозерский р-н, Ленинградская обл.), садки которого установлены в оз. Суходольское. Дикие и заводские особи имеют общее происхождение – маточное стадо сформировано из потомства, полученного от рыб, обитающих в естественном водоеме.
В «летний» сезон (с мая по октябрь) судаки в садках получали искусственные корма Biomar Efico Sigma 840 (Дания). В зимний период (с конца октября по май) судака в садках не кормили. Рыб из озера для исследований отлавливали 15–17 октября 2021 г. и 10–15 мая 2022 г., из садков – 25 октября 2021 г. и 15 мая 2022 г. Температурный режим воды в период зимовки судаков в озере (и, соответственно, в садках) приведен на графике (рис.).
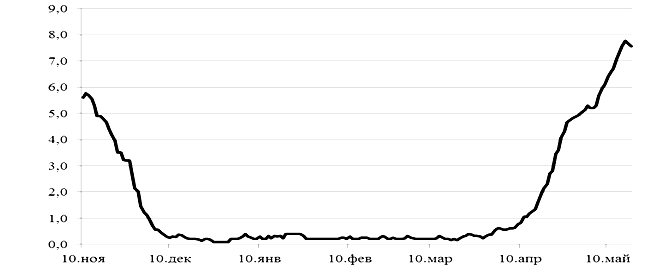
Динамика температуры воды во время зимовки судака в садках и оз. Суходольское (2021–2022 гг.)
Dynamics of water temperature during the wintering of sander in cages and Lake Sukhodolskoye (2021-2022)
Аналитическую работу проводили в лаборатории аквакультуры Санкт-Петербургского филиала Всероссийского научно-исследовательского института рыбного хозяйства и океанографии, жирнокислотный состав мышц и печени рыб определяли в ООО «МИП-АМТ» (г. Санкт-Петербург). Количество исследованных рыб из каждой группы для морфофизиологического анализа составляло по 15 экз., для химического анализа – по 10 экз., для жирнокислотного анализа состава мышц и печени – по 5 экз.
Длину рыб определяли до конца чешуйного покрова, индексы внутренних органов (печени, гонад, сердца, селезенки, желудочно-кишечного тракта (ЖКТ)) и полостного жира определяли как процентное отношение их массы к массе целой рыбы. Коэффициент упитанности рассчитывали как отношение массы к длине тела рыбы до конца чешуйного покрова, возведенной в куб.
Методами химического анализа определяли относительное содержание влаги, сухого вещества, липидов (по методу Фолча), белка (по методу Кьельдаля), минеральных веществ (золы) – методом сжигания пробы в муфельной печи при температуре 550 °С до постоянной массы золы, безазотистых экстрактивных веществ (БЭВ) – расчетным методом, витамина С – модифицированным методом титрования экстракта витамина в соляной кислоте реактивом Тильманса [6]. Для определения данных показателей из нескольких образцов рыб готовили интегральную пробу. Анализ жирнокислотного состава тела судаков проводили методом газожидкостной хроматографии.
Статистическая обработка данных была выполнена с помощью программного пакета Statisica 6.0. В таблицах приведены среднеарифметические значения (M), стандартная ошибка среднего (m), коэффициент вариабельности признака (Cv). Для определения различий между группами использовали t-критерий Стьюдента, нормальность распределения определяли по критерию Пирсона.
Результаты исследования
Размерно-весовые характеристики судаков, обитающих в природном водоеме и содержащихся в садках рыбоводного хозяйства, за период зимовки достоверно не изменились. Тем не менее и у диких, и у заводских рыб произошло значимое снижение коэффициента упитанности – на 7 и 11 % соответственно (табл. 1).
Таблица 1
Table 1
Размерно-весовые показатели и индексы внутренних органов «диких»
и «заводских» судаков до и после зимовки
Size and weight indicators and indices of internal organs of “wild”
and farm sander before and after wintering
|
Показатель |
Дикие особи |
Заводские особи |
||||||
|
Осень |
Cv |
Весна |
Cv |
Осень |
Cv |
Весна |
Cv |
|
|
Длина, см |
27,6 ± 0,4 |
4,2 |
28,5 ± 0,5 |
2,71 |
26,2 ± 0,5 |
6,22 |
28,8 ± 0,6 |
6,31 |
|
Масса, г |
278,4 ± 14,0 |
11,8 |
284,0 ± 11,0 |
8,70 |
242,1 ± 16,2 |
21,20 |
264,8 ± 19,2 |
22,93 |
|
Коэффициент |
1,33 ± 0,03 |
6,50 |
1,23 ± 0,03* |
2,57 |
1,25 ± 0,01 |
3,79 |
1,11 ± 0,01* |
4,39 |
|
Индекс, % |
||||||||
|
Полостного жира |
1,87 ± 0,25 |
42,07 |
1,04 ± 0,18* |
39,19 |
8,61 ± 0,29 |
10,75 |
7,24 ± 0,37* |
15,97 |
|
Печени |
1,42 ± 0,10 |
21,39 |
1,51 ± 0,10 |
14,31 |
3,68 ± 0,21 |
17,80 |
3,49 ± 0,12 |
10,88 |
|
Сердца |
0,20 ± 0,02 |
27,56 |
0,20 ± 0,01 |
24,50 |
0,15 ± 0,01 |
19,24 |
0,14 ± 0,01 |
22,58 |
|
Селезенки |
0,11 ± 0,01 |
21,14 |
0,09 ± 0,01 |
12,38 |
0,09 ± 0,01 |
26,58 |
0,11 ± 0,01 |
42,89 |
|
Гонадосоматический |
0,15 ± 0,04 |
78,87 |
0,35 ± 0,11* |
70,93 |
0,73 ± 0,21 |
88,82 |
1,14 ± 0,09* |
58,88 |
|
ЖКТ |
4,29 ± 0,40 |
29,44 |
3,89 ± 0,17* |
9,76 |
2,82 ± 0,14 |
15,29 |
2,78 ± 0,10 |
11,51 |
* Достоверные различия признаков в каждой группе при уровне значимости p ≤ 0,05.
За зимовку у судаков достоверно снизились индексы полостного жира – на 45 % у диких рыб и на 16 % у заводских, повысились гонадосоматические индексы (ГСИ) – в среднем на 47 %, и на 9 % снизился индекс ЖКТ у судаков из естественных условий обитания.
Изменения химического состава у диких рыб за зимний период заключались в достоверном снижении липидов в мышцах и печени на 26 и 59 % соответственно, белка в печени на 17 %, витамина С в мышцах на 56 %, а также в повышении БЭВ в мышцах и печени на 47 и 31 % соответственно и золы в печени на 15 % (табл. 2).
Таблица 2
Table 2
Химический состав мышц и печени «диких» и «заводских» судаков до и после зимовки
(липиды, белок, зола и БЭВ определены в сыром веществе), %
Chemical composition of muscles and liver of “wild” and farm sander before and after wintering
(lipids, protein, ash and NFES are determined in raw matter), %
|
Компонент |
Дикие особи |
Заводские особи |
||||||
|
Осень |
Cv |
Весна |
Cv |
Осень |
Cv |
Весна |
Cv |
|
|
Мышцы |
||||||||
|
Влага |
77,78 ± 0,21 |
0,47 |
76,73 ± 0,08 |
0,18 |
75,39 ± 0,20 |
0,46 |
75,90 ± 0,10 |
0,29 |
|
Сухое вещество |
22,22 ± 0,21 |
1,65 |
23,27 ± 0,08 |
0,60 |
24,61 ± 0,20 |
1,41 |
24,10 ± 0,10 |
0,91 |
|
Липиды |
0,84 ± 0,18 |
60,3 |
0,62 ± 0,02* |
5,63 |
0,85 ± 0,04 |
8,64 |
0,91 ± 0,05 |
12,47 |
|
Белок |
19,00 ± 0,47 |
4,26 |
19,15 ± 0,19 |
1,70 |
21,01 ± 0,21 |
1,75 |
19,87 ± 0,13 |
1,46 |
|
Зола |
1,30 ± 0,03 |
3,86 |
1,21 ± 0,01 |
0,48 |
1,42 ± 0,01 |
0,70 |
1,38 ± 0,02 |
2,73 |
|
БЭВ |
1,08 ± 0,22 |
34,65 |
2,29 ± 0,09* |
6,81 |
1,33 ± 0,03 |
4,15 |
1,94 ± 0,09* |
10,51 |
|
Витамин С, мкг/г |
28,43 ± 0,18 |
1,07 |
21,60 ± 0,23* |
1,85 |
39,33 ± 1,44 |
6,36 |
30,42 ± 0,48* |
3,54 |
|
Печень |
||||||||
|
Влага |
69,26 ± 1,08 |
2,71 |
71,76 ± 0,48 |
1,16 |
51,72 ± 2,39 |
7,99 |
51,49 ± 1,58 |
6,87 |
|
Сухое вещество |
30,74 ± 1,08 |
6,12 |
28,24 ± 0,48 |
2,94 |
48,28 ± 2,39 |
8,56 |
48,51 ± 1,58 |
7,30 |
|
Липиды |
11,09 ± 1,10 |
17,21 |
4,49 ± 0,22* |
8,35 |
26,58 ± 1,98 |
12,90 |
22,31 ± 1,48* |
14,83 |
|
Белок |
16,63 ± 1,38 |
14,41 |
13,75 ± 0,51* |
6,40 |
11,21 ± 0,24 |
3,70 |
11,53 ± 0,36 |
7,04 |
|
Зола |
1,11 ± 0,04 |
5,98 |
1,31 ± 0,08* |
11,10 |
0,85 ± 0,04 |
7,98 |
1,10 ± 0,03* |
6,20 |
|
БЭВ |
3,14 ± 0,17 |
9,59 |
8,69 ± 0,33* |
6,56 |
9,64 ± 1,09 |
19,54 |
13,57 ± 0,57* |
9,43 |
|
Витамин С, мкг/г |
87,90 ± 2,77 |
5,46 |
86,50 ± 0,72 |
1,45 |
67,50 ± 9,20 |
23,61 |
66,92 ± 4,02 |
13,42 |
* Достоверные различия признаков в каждой группе при уровне значимости p ≤ 0,05.
У заводских рыб также достоверно снизилось содержание липидов в печени (на 16 %) и витамина С в мышцах (на 23 %) и увеличилась зольность печени (на 23 %) и БЭВ в мышцах и печени (на 31 и 29 %) соответственно.
У диких рыб наибольшей вариабельностью в осенний и весенний период отличается индекс полостного жира (около 40 %), содержание липидов в мышцах в осенний период (60 %) и ГСИ (не менее 70 %) (см. табл. 1, 2). Судакам из садков также свойственна значительная вариабельность ГСИ, которая несколько снижается по мере созревания рыб – от 89 % осенью до 59 % весной (см. табл. 1).
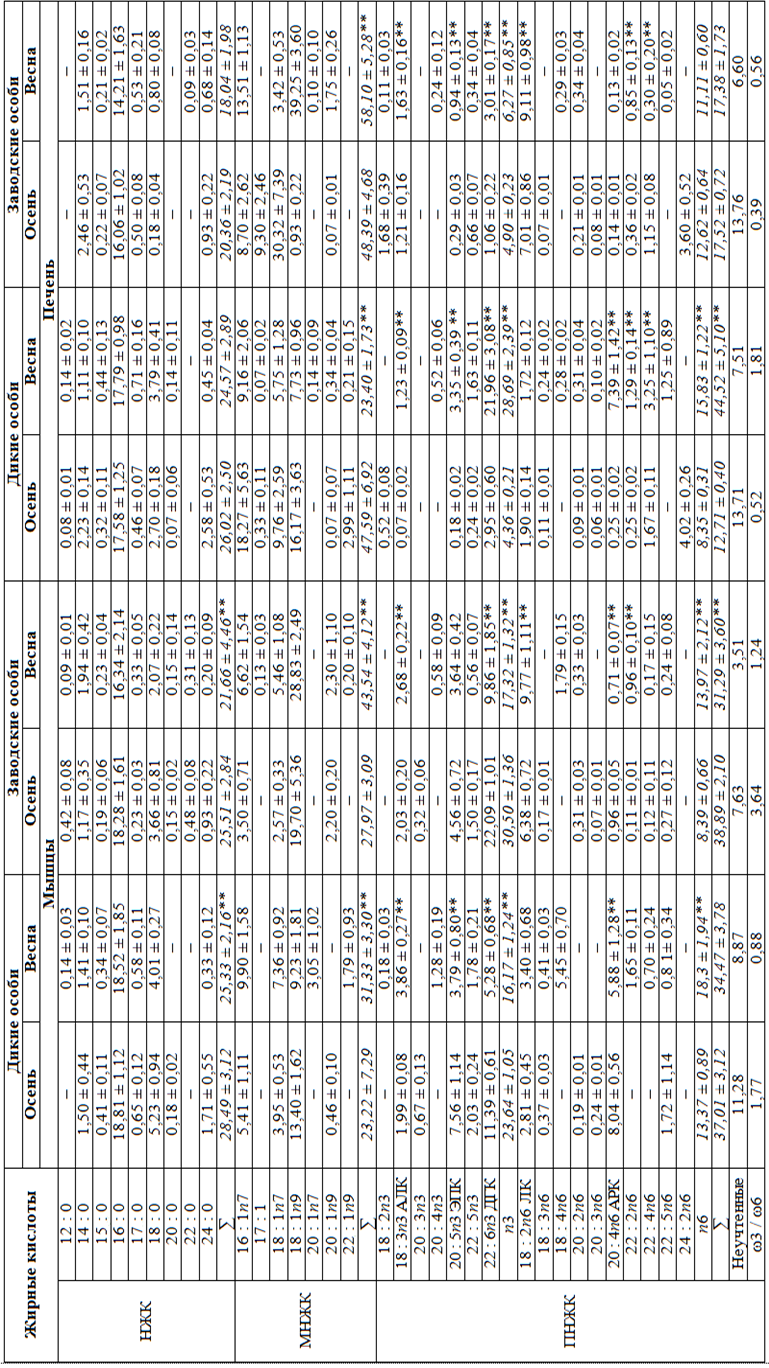
Жирнокислотный состав липидов мышц у судаков из озера и садков после зимовки характеризовался снижением насыщенных (НЖК) и полиненасыщенных (ПНЖК) и повышением мононенасыщенных жирных кислот (МНЖК), при этом существенно снизилось содержание кислот ряда n3 и повысилось n6 (табл. 3).
Содержание доли наиболее значимой в физиологическом отношении докозагексаеновой (ДГК) жирной кислоты снизилось более чем в 2 раза, эйкозапентаеновой (ЭПК) – в среднем в 1,6 раза на фоне повышения в 1,6 раза материнской α-линоленовой (АЛК) жирной кислоты. Незаменимая арахидоновая жирная кислота (АРК) за зимовку судаков разного происхождения снизилась в липидах мышц в среднем в 1,4 раза, и во столько же раз в среднем увеличилось содержание материнской линолевой (ЛК) жирной кислоты.
Доля МНЖК в печени диких рыб за зимовку снизилась в 2 раза при более чем трехкратном повышении ПНЖК. У заводских рыб, напротив, доля МНЖК повысилась в 1,2 раза, ПНЖК – существенно не изменилась (см. табл. 3).
Увеличение ПНЖК в липидах печени рыб из естественного водоема в зимний период произошло за счет повышения доли кислот ряда n3 (в 6,6 раза) – доля АЛК повысилась в 2,8 раза, ЭПК в 18,6, ДГК в 7,4 раза. Также произошло двукратное увеличение содержания n6 кислот, в основном за счет АРК, доля которой повысилась с 0,25 до 7,39 % от суммы ЖК (т. е. практически в 30 раз), при этом материнская ЛК осталась неизменной (см. табл. 3).
Несмотря на стабильный уровень ПНЖК в липидах печени заводских рыб за период зимнего содержания, структура их жирнокислотного состава несколько изменилась. Так, произошло увеличение в 1,3 раза доли кислот ряда n3 за счет повышения АЛК в 1,4 раза, ЭПК и ДГК в 3 раза каждой. Суммарная доля кислот семейства n6, в том числе АРК, у заводских рыб за зимовку осталась на прежнем уровне, достоверно увеличились доли линолевой
и докозадиеновой кислот и снизилась доля докозатетраеновой жирной кислоты (см. табл. 3).
Содержание наиболее значимых для питания человека ПНЖК в мышцах рыб (в г/100 г) указывает на снижение питательной ценности мяса судака после зимовки (табл. 4).
Таблица 4
Table 4
Содержание незаменимых в питании человека полиненасыщенных ЖК в мышцах судака, г/100 г
The content of polyunsaturated fatty acids in human nutrition in pike perch muscles, g/100 g
|
Жирные кислоты |
Дикие особи |
Заводские особи |
||
|
Осень |
Весна |
Осень |
Весна |
|
|
20 : 5n3 ЭПК |
0,06 ± 0,01 |
0,02 ± 0,003* |
0,04 ± 0,003 |
0,03 ± 0,001 |
|
22 : 6n3 ДГК |
0,10 ± 0,01 |
0,03 ± 0,003* |
0,19 ± 0,02 |
0,09 ± 0,01* |
|
ЭПК + ДГК |
0,16 ± 0,02 |
0,05 ± 0,004* |
0,23 ± 0,02 |
0,12 ± 0,01* |
|
∑ n3 |
0,20 ± 0,02 |
0,10 ± 0,01* |
0,26 ± 0,02 |
0,16 ± 0,02* |
|
20 : 4n6 АРК |
0,07 ± 0,01 |
0,04 ± 0,002 |
0,01 ± 0,001 |
0,01 ± 0,001 |
|
∑ n6 |
0,11 ± 0,01 |
0,11 ± 0,01 |
0,07 ± 0,01 |
0,13 ± 0,01* |
|
∑ ПНЖК |
0,31 ± 0,02 |
0,21 ± 0,02* |
0,33 ± 0,02 |
0,28 ± 0,02* |
|
n3 / n6 |
1,8 ± 0,04 |
0,9 ± 0,01* |
3,7 ± 072 |
1,2 ± 0,08* |
* Достоверные различия признаков в каждой группе при уровне значимости p ≤ 0,05.
Как у диких, так и у заводских рыб наблюдается достоверное снижение ЭПК, ДГК и их сумм, а также АРК и общего содержания кислот ряда n3
и ПНЖК в целом. Стабильным остается содержание в мышцах диких рыб кислот семейства n6, у заводских рыб содержание этих кислот в мышцах за зимовку увеличивается примерно в 2 раза. Также значительно снижается величина отношения n3 / n6 жирных кислот в липидах мышц судака после зимовки – в 2 раза у диких рыб и в 3 раза у заводских (см. табл. 4).
Обсуждение
Снижение коэффициента упитанности рыб разного происхождения в период зимовки связано в первую очередь с расходованием полостного жира, а также жира мышц (только у диких судаков) и печени, что подтверждается данными морфофизиологических и химических исследований (см. табл. 1, 2) и согласуется с литературными данными [7, 8]. На фоне снижения липидов в мышцах и печени судаков значительно повышается содержание углеводов (БЭВ), которые активно синтезируются у разных видов рыб с понижением температуры воды и в зимний период депонируются в мышечной ткани и печени в виде гликогена [9]. Напротив, отсутствие возможности у костистых рыб в период голодания обеспечить потребности организма в витамине С за счет собственного синтеза [10] приводит к его достоверному снижению в мышцах диких и заводских рыб. При этом уровень витамина в печени за зимовку не претерпел изменений, чему, вероятно, может способствовать транспорт витамина С из мышц в значительно более жирную печень (см. табл. 2), в том числе с целью предотвращения в ней перекисного окисления липидов. На важную роль витамина С в противоокислительных процессах в печени и сохранении длинноцепочечных эссенциальных жирных кислот мы обращали внимание ранее, в исследовании сиговых видов рыб [11].
Уменьшение за зимовку размеров ЖКТ у диких особей связано со значительным снижением активности питания – в период нагула судак способен потреблять рыбу, достигающую в размерах до 50 % от его собственной длины [12], однако при снижении температуры воды до 4,5 °C и ниже судак практически не питается [13]. Для каспийского судака в период нагула установлена возможность сильного растяжения стенок желудка с изменением его морфофизиологических параметров и восстановления органа в зимнее время [14]. У заводских рыб, напротив, пищей служат искусственные корма, гранулы которых, в силу своих сравнительно небольших размеров и удобоваримости, не способствуют развитию массивного ЖКТ [15], и он в период зимовки не претерпевает существенных изменений.
С неодинаковой интенсивностью питания судака в естественных условиях, а также доступностью корма и его питательной ценностью можно связать значительную вариабельность в содержании липидов в теле диких рыб (полостной жир и жирность мышц). Вариабельность ГСИ у судаков из естественных условий и садков для данного вида можно считать нормой – известно, что развитие ооцитов трофоплазматического роста у многих видов рыб в природе синхронизируется к моменту наступления IV стадии зрелости [16, 17]. Исследованные нами особи находились на III стадии развития гонад.
Основные изменения в жирнокислотном статусе судака в процессе зимовки характеризуются расходованием в мышцах рыб наиболее функциональных длинноцепочечных ПНЖК – ДГК, ЭПК и АРК, что может быть связано с адаптацией организма к низкотемпературным условиям. Известно, что одним из основных механизмов биохимической адаптации гидробионтов к низким температурам является увеличение степени ненасыщенности ЖК [18, 19]. Так, во многих исследованиях сообщалось, что у рыб в летние месяцы и у рыб из теплых регионов в липидах, как правило, преобладают НЖК и МНЖК, тогда как присутствие в больших количествах ПНЖК характерно для рыб северных широт и в зимний холодноводный период [20–22].
Другой причиной снижения ПНЖК в мышцах судака с осени по весну может являться созревание рыб [8], в процессе которого происходит метаболизм и депонирование указанных выше жирных кислот в печени с последующей транспортировкой их и других питательных веществ в гонады [23].
В наших исследованиях у диких судаков за зимовку в печени повысились ЭПК в 18,6 раза, ДГК в 7,4 раза, АРК в 30 раз. На снижение доли ПНЖК в мышцах судака и их накопление в печени под влиянием созревания косвенно указывают достоверное повышение ГСИ у рыб из озера (в 2,3 раза) и садков рыбоводного хозяйства (в 1,6 раза) и переход гонад в стадию трофоплазматического роста (в большей степени это свойственно заводским рыбам).
Схожая тенденция в метаболизме длинноцепочечных ПНЖК наблюдалась в период созревания у других видов рыб, в том числе морских. Так, у самок японской бородавчатки Inimicus japonicas при созревании гонад от III к V стадии содержание ЭПК, ДГК и АРА в мышцах снижалось, тогда как в печени двукратно увеличивалось [24]. При этом в гонадах достоверных изменений в содержании основных ПНЖК во время созревания самок не происходило. Есть сведения о стабильном содержании ЭПК и ДГК в яичниках речного окуня Perca fluviatilis на последних стадиях репродуктивного цикла, в процессе которого происходило снижение указанных кислот в мышцах и печени [25].
Примечательно, что существенное расходование ПНЖК в мышцах судака за зимовку наблюдается как в северной части его ареала (настоящие исследования), так и на южной границе – в оз. Эгирдир в Турции [26], где у судака происходит снижение доли ПНЖК в липидах мышц с ноября (среднемесячная температура 9,8 °C) по май (18,1 °C) с 38,5 по 12,1 % от суммы ЖК, при этом доля ДГК снижается с 16,3 до 0,7 % от суммы ЖК.
Известно, что понижение доли ПНЖК в тканях рыб сопровождается адаптивным процессом – увеличением МНЖК, которые рыбы способны синтезировать с использованием НЖК для поддержания текучести жира и проницаемости мембран клеток в условиях низких температур водной среды [11]. Это подтверждается результатами настоящих исследований: у дикого и заводского судака за зимовку произошло повышение МНЖК в липидах мышц на 26–36 %, в печени заводских рыб – снижение на 17 %. У диких особей, напротив, произошло увеличение МНЖК в печени на фоне понижения НЖК на 11–15 % в мышцах и 6–11 % в печени.
Безусловно, подобные сезонные изменения жирнокислотного состава липидов мышц отражаются на питательной ценности судака, индикатором которой принято считать сумму содержания в мышцах рыб ЭПК + ДГК [27, 28]. Исходя из данных табл. 4, для удовлетворения суточной потребности человека в ЭПК и ДГК, равной 0,5–1,0 г (рекомендации Всемирной организации здравоохранения [29]), в сутки необходимо потреблять не менее 300 г филе дикого или чуть более 200 г филе искусственно выращенного судака, пойманного в осенний период, и 1 кг филе дикой или 400 г заводской рыбы, выловленной поздней весной.
На более высокую пищевую ценность филе осенних рыб указывают также соотношения n3 / n6 ЖК, которые в 2–3 раза выше таковых значений
у судаков, отловленных весной, и составляют 1,8 у диких и 3,7 у заводских рыб. Тем временем для питания человека оптимальным является соотношение n3 / n6 ПНЖК в пропорции 1 : 1 или 1 : 2 [30]. Учитывая сведения того же автора о смещении в пище баланса n3 / n6 в сторону значительного увеличения n6 (до 1 : 20–25), можно предположить, что использование аквакультурной продукции с повышенным содержанием длинноцепочечных n3 ПНЖК будет способствовать сбалансированности современного рациона человека по незаменимым жирным кислотам.
Таким образом, можно заключить, что в осенний период филе как дикого, так и выращенного на искусственных кормах судака является более ценным для человека продуктом по содержанию незаменимых жирных кислот, чем филе судака после зимовки, что соответствует литературным данным [8].
Заключение
За период зимовки у судаков, выловленных из естественной среды обитания и выращенных в заводских условиях, происходят изменения в морфофизиологическом и химическом статусе, направленные на расходование энергетических резервов организма и перераспределение питательных веществ в мышцах и печени. Это выражается в значительном снижении содержания полостного жира, жирности мышц и печени, витамина С в мышцах. При этом в мышцах и печени многократно повышается БЭВ. В мышцах судаков в зимний период существенно повышается уровень МНЖК и снижается уровень ПНЖК, особенно ЭПК, ДГК, АРК, на фоне повышения уровня этих кислот в печени. Подобные изменения являются адаптацией рыб к холодным температурным условиям в период длительной зимовки, а также связаны с развитием гонад.
С точки зрения качества филе, оцененного по содержанию длинноцепочечных ПНЖК, у выловленных и выращенных рыб за зимовку происходит значительное снижение их пищевой ценности, что выражается в 2–3-кратном снижении ЭПК + ДГК. Следовательно, для рациона человека наиболее полезным будет дикий или аквакультурный судак, выловленный осенью.
Результаты настоящей работы определяют необходимость дальнейших физиолого-биохимических исследований диких и культивируемых рыб с учетом сезонных изменений условий их обитания, определяющих ход адаптивных и физиологических процессов, направленных на нормализацию гомеостаза в условиях экстремально низких температур (до 0,1 °C). Кроме того, исследование питательной ценности рыб в зависимости от сезона выращивания может способствовать рациональному ведению рыбного хозяйства и получать рыбоводную продукцию с высокой питательной ценностью.
1. Гримм О. А. Обзор деятельности Никольского рыбоводного завода за 50 лет его существования. СПб.: Департамент земледелия, 1905. 34 с. URL: https://elib.rgo.ru/safe-view/123456789/212360/1/UnVQUkxJQjEyMDg0MzEyLlBERg==#1 (дата обращения: 23.03.2023).
2. Хрусталев Е. И., Дельмухаметов А. Б. Техноло-гия формирования маточного стада судака в установках с замкнутым циклом водообеспечения // Рыб. хоз. 2012. № 1. С. 70-72.
3. Лютиков А. А., Королев А. Е., Шумилина А. К., Вылка М. М. Опыт выращивания ремонтно-маточного стада судака Sander lucioperca в садках в Ленинградской области // Водные биологические ресурсы России: состояние, мониторинг, управление: II Всерос. науч. конф., посвящен. 90-летию КамчатНИРО (Петропавловск-Камчатский, Россия, 04-05 апреля 2022 г.): сб. материалов конф. Петропавловск-Камчатский: Изд-во КамНИРО, 2022. С. 219-226. DOI:https://doi.org/10.15853/978-5-902210-44-3.
4. Ben Ammar I., Teletchea F., Milla S., Ndiaye W. N., Ledore Y., Missaoui H., Fontaine P. Continuous lighting inhibits the oneset of reproductive cycle in pikeperch // Fish Physiol. Biochem. 2015. V. 41. P. 344-356.
5. Khendek A., Chakraborty A., Roche J., Ledore Y., Personne A., Policar T., Żarski D., Mandiki R., Kestemont P., Milla S. Rearing conditions and life history influence the progress of gametogenesis and reproduction performances in pikeperch males and females // Animal. 2018. V. 12. P. 2335-2346.
6. Князева Л. М. Рекомендации по увеличению сроков хранения гранулированного корма для молоди форели путем опрыскивания его водным раствором витамина С. Л.: Изд-во ГосНИОРХ, 1979. 12 с.
7. Kirjasniemi M., Valtonen T. Winter mortality of young of-the-year pikeperch (Stizostedion lucioperca) // Ecol. Freshwater Fish. 1997. V. 6. P. 155-160.
8. Uysal K., Aksoylar M. Y. Seasonal variations in fatty acid composition and the n-6/n-3 fatty acid ratio of pikeperch (Sander lucioperca) muscle lipids // Ecology of Food and Nutrition. 2005. V. 44. P. 23-35.
9. Love R. M. The chemical biology of fishes. London: Academic Press, 1980. V. 2. 943 p.
10. Ching B., Chew S. F., Ip Y. K. Ascorbate synthesis in fishes: a review // IUBMB Life. 2015. V. 67. P. 69-76.
11. Остроумова И. Н., Костюничев В. В., Лютиков А. А., Шумилина А. К., Филатова Т. А. Влияние витамина C на жирнокислотный состав печени и мышц двухлеток сиговых рыб (Coregonidae), выращиваемых в условиях аквакультуры // Вопр. рыболовства. 2020. Т. 21. № 3. С. 343-352.
12. Keskinen T. Feeding ecology and behaviour of pikeperch, Sander lucioperca (L.), in boreal lakes // Jyväskylä Studies in Biological and Environmental Studies. Jyväskylä: Jyväskylä University Printing House, 2008. 54 p.
13. Malinovskyi O., Blecha M., Křišťan J., Policar T. Feeding activity of pikeperch (Sander lucioperca) under winter and spring photothermal conditions of pre-spawning period // Aquac. Res. 2019. P. 1-6. https://doi.org/10.1111/are.14401.
14. Устарбеков А. К., Курбанов З. М., Магомедов Т. А. Строение и сезонные изменения микроструктуры пищеварительного тракта обыкновенного судака Stizostedion lucioperca (Linnaeus, 1758) в связи с характером питания в условиях дагестанского побережья Каспия // Изв. ДГПУ. 2010. № 1. С. 58-65.
15. Лютиков А. А., Королев А. Е. Сравнительный морфометрический и морфологический анализ судаков Sander lucioperca из прудов, озера и выращенных в индустриальных условиях // Актуальные вопросы пресноводной аквакультуры. М.: Изд-во «ИП Сорокин», 2022. Вып. 93. С. 89-95.
16. Дрягин П. А. Половые циклы и нерест рыбы // Изв. ВНИОРХ. 1949. Т. 28. С. 3-113.
17. Кошелев Б. В. Экология размножения рыб. М.: Наука, 1984. 309 с.
18. Cossins A. R., Prosser C. L. Evolutionary adapta-tion of membranes to temperature // Proceedings of National Academy of Sciences of USA. 1978. V. 75. P. 2040-2043.
19. Falk-Petersen S., Hopkins C. E., Sargent J. R. Trophic relationships in the pelagic, arctic food web // Trophic relationships in the Marine Environment. Proceed-ings of the 24-th European Marine Biology Symposium. Aberdeen: Aberdeen University Press, 1990. P. 315-333.
20. Dutta H., Das A., Farkas T. The role of environmental temperature in seasonal changes of fatty acid composition of hepatic lipid in an air-breathing Indian Teleost (Channa punctatus) // Comparative Biochemistry and Physiology. 1985. V. 81. P. 341-347.
21. Skuladottir G. V., Schioth H. B., Gudmundsdottir B., Richards B., Jonsson L. Fatty acid composition of muscle heard and liver lipids in Atlantic Salmon, Salmo Salar, at extremely low environmental temperature // Aquaculture. 1990. V. 84. P. 71-80.
22. Dey I., Buda C., Wiik H., Halver J. E., Farkas T. Molecular and structural composition of phospholipid membranes in livers of marine and freshwater fish in relation to temperate // Proceeding of National Academy of Sciences of USA. 1993. V. 90. P. 7498-7502.
23. Шумилина А. К. Физиологическая характеристика производителей пеляди, выращиваемых в индустриальных условиях, и их пищевые потребности // Сб. науч. тр. ГосНИОРХ. 2005. Вып. 333. С. 60-114.
24. Songlin L., Wen W., Xu G., Xuxiong H., Naisong C. Variation of lipids and fatty acids composition in the tissues of wild devil stinger (Inimicus japonicas) during sexual maturation // Aquaculture and Fisheries. 2018. V. 3. P. 115-121.
25. Рудченко А. Е. Роль трофических факторов в формировании жирнокислотного состава рыб, обитающих в водоемах красноярского края: автореф. дис. … канд. биол. наук. Красноярск: Изд-во СФУ, 2018. 24 с.
26. Çağlak E., Karsli B. Seasonal variation of fatty acid and amino acid compositions in the muscle tissue of zander (Sander lucioperca Linnaeus, 1758) and the evaluation of important indexes related to human health // Ital. J. Food Sci. 2017. V. 29. P. 266-275.
27. Kris-Etherton P. M., Harris W. S., Appel L. J. Fish consumption, fish oil, omega-3 fatty acids, and cardiovascular disease // Circulation. 2002. V. 106. P. 2747-2757.
28. Adkins Y., Kelley D. S. Mechanisms underlying the cardioprotective effects of omega-3 polyunsaturated fatty acids // J. Nutr. Biochem. 2010. V. 21. P. 781-792.
29. Гладышев М. И. Наземные источники полиненасыщенных жирных кислот для аквакультуры // Вопр. ихтиологии. 2021. Т. 61, № 4. С. 471-485.
30. Simopoulos A. P. Human requirement for n-3 polyunsaturated fatty acids // Poultry Sci. 2000. V. 79. P. 961-970.
