











Россия
Россия
Исследован рост обыкновенного (речного) окуня в водоемах из системы р. Ишим в пределах Республики Казахстан. Приводятся данные по 29 выборкам из различных водоемов Ишимского и некоторых смежных бассейнов. Наиболее протяженный возрастной ряд имели особи из оз. Лобаново (национальный парк «Кок-шетау»): шестнадцатый год (15+). Речные популяции отличались менее продолжительным сроком жизни. Сравнение картин темпов роста демонстрирует наличие двух условных «юнитов»: первый объединяет большинство озерных популяций, второй – вдхр. Каратомарское и оз. Лобаново. Часть выборок располагается вне этих объединений. Исследованные хронологические выборки частью имеют достаточно существенные различия в росте. Это характерно для оз. Караколь и, отчасти, вдхр. Ишимского, где в промежутках между отловами окуня произошли позитивные гидрологические изменения. Это стало причиной увеличения темпов роста. Для этих выборок характерен низкий уровень корреляции показателей роста между смежными возрастами, что показывает нарушение «канализации» темпов роста и обретения возможностей для индивидуумов в обновленной разбалансированной среде обитания. В относительно стабильных условиях большинства прочих водоемов уровень корреляции данных показателей высок по большинству пар сравнения. При сравнении ишимских популяций с ранее опубликованными данными по бассейну рек Нура и Сарысу по темпам роста эмпирически можно сделать заключение о более успешном росте окуня в Нура-Сарысуйском бассейне. Однако следует учитывать, что в Ишимском бассейне были изучены в основном водоемы, на которых отсутствует промысловый лов и развито рекреационное рыболовство. В противоположность этому в Нура-Сарысуйской системе были обследованы водоемы с преобладающим промысловым ловом. Различное воздействие этих типов лова, вероятно, определяет и темпы роста, и продолжительность жизни окуня.
окунь, Ишим, возраст, водоем, рост, прирост длины тела, выборка
Введение
Река Ишим (Есиль) начинается в горах Нияз в Карагандинской области Республики Казахстан и впадает в р. Иртыш в Омской области Российской Федерации. Общая протяженность реки – 2 450 км, по территории Казахстана – 1 607 км. Площадь всего бассейна составляет 177 тыс. км2 [1].
Ихтиофауна системы сложена обычными видами, характерными для Обь-Иртышского бассейна, характеризуется полным отсутствием эндемизма [2]. Наиболее обильными и широко распространенными видами являются караси серебряный Carassius gibelio (Bloch, 1782), золотой C. carassius (L., 1758), плотва Rutilus lacustris (Pallas, 1814), елец Leuciscus baikalen
sis (Dybowsky, 1874) и окунь Perca fluviatilis L., 1758.
Окунь как массовый и эвритопный вид широко распространен по бассейну от верховьев реки до ее устья и населяет различные типы водоемов, избегая сильно эвтрофированных.
Данная работа посвящена анализу роста особей окуня в различных водоемах Ишимского бассейна в пределах Республики Казахстан. Были исследованы различные аспекты межгрупповой изменчивости роста.
Материалы и методы
Работа выполнялась в 2012–2023 гг. Были исследованы 29 водоемов (участков реки) в бассейне р. Ишим и 4 – из соседних водных систем: 2 рукава вдхр. Каратомарского (бас. р. Тобол), вдхр. Кояндинское (бас. р. Силеты) и оз. Караколь из ишимско-олентинского междуречья. Всего было проанализировано 2 400 экз. окуня. Расширенный анализ роста был проведен по 17 популяциям окуня, включающим 21 выборки, 4 – повторные, взятые через промежуток, больший продолжительности жизни 1 поколения (3 года для окуня). Материалы по 12 малочисленным выборкам приведены для сведения. Точки отлова показаны на рис. 1.
Для определения возраста и обратного расчисления роста использовалась operculum [3], согласно нашим предшествующим наработкам [4]. Обратное расчисление роста проводилось методом простых пропорций Даля-Лея (Dahl-Lea), согласно [5].
Натуральные приросты GNi определялись по следующей формуле [6, 7]:
GNi = li – li–1,
где l – рассчитанная длина тела в какой-либо год; i – временной показатель (год).
На основании натуральных приростов рассчитывались удельные приросты GS в диапазоне до 6 лет включительно [4]:
GS = (GNi / l6) ∙ 100 %.
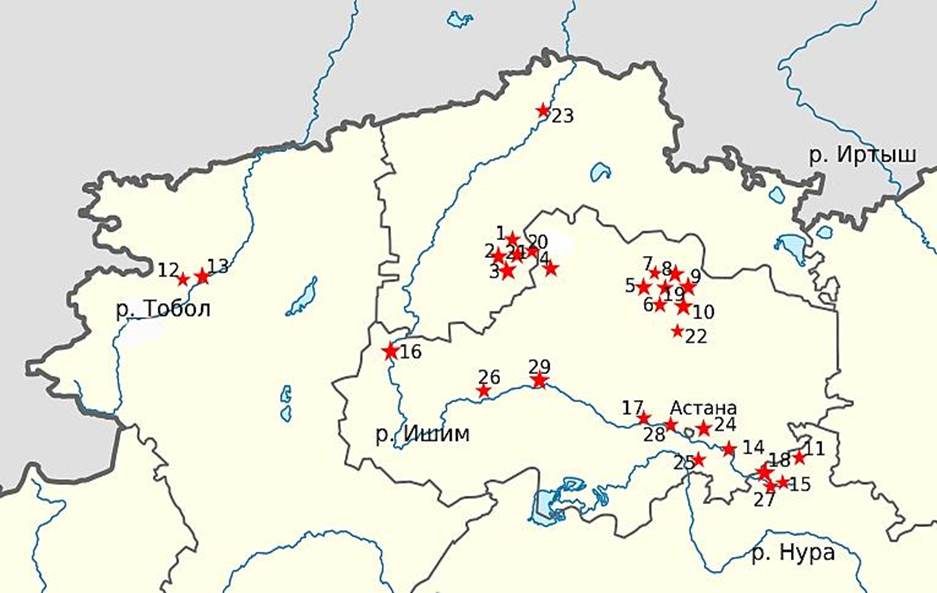
Рис. 1. Карта расположения исследованных популяций окуня (нумерация точек отлова соответствует табл. 2)
Fig. 1. A map of the location of the studied perch populations (the numbering of the capture points corresponds to Table 2)
На основании полученных данных рассчитывались меры сходства-различия и строились дендрограммы. Практически все методические подходы давали при этом идентичные или близкие результаты. Для иллюстрации сходства в данном исследовании были выбраны 2 варианта: межгрупповой связи с использованием квадрата евклидова расстояния и медианной кластеризации с использованием того же квадрата евклидова расстояния. Выборки разделялись методом иерархической кластеризации [8, 9].
Коэффициент корреляции Пирсона и иные статистические индикаторы вычислялись по стандартным методикам [10] с использованием программ MS Excel 2003 [11] и IBM SSPS Statistics v. 22 [12].
Были приняты следующие сокращения и обозначения: F – значение F-критерия, α – уровень достоверности.
Результаты
Изученные популяции населяют разнотипные водоемы в системе Ишима и сопредельных бассейнов. Гидроморфологические и гидрохимические показатели водоемов приведены в табл. 1.
В основном эти водоемы представлены стоячими или слабопроточными водоемами. Даже сама р. Ишим представлена участками с замедленным течением.
Многие из исследованных водоемов обладают значительной по региональным меркам площадью и другими линейными параметрами. Однако по-настоящему глубоких водоемов со средними глубинами более 7 м не так уж и много: оз. Щучье, Б. Чебачье и водохранилище Вячеславское.
Большинство водоемов имеет минерализацию, близкую к 1 г/л. Повышенная минерализация характерна только для оз. Шалкар и М. Чебачье. Водоемов с соленостью ниже 0,5 г/л также немного. Преобладающие анионы – хлориды и гидрокарбонаты, катионы – натрий, изредка – кальций. Водородный показатель колеблется от 7,35 до 8,88, т. е. вода имеет нейтральную либо слабощелочную реакцию. Вода в основном мягкая или умеренно жесткая, за исключением двух минерализованных озер, где жесткость высокая.
Обратное расчисление роста окуня в исследованных водоемах дано в табл. 2.
Таблица 1
Table 1
Характеристика основных исследованных водоемов
Characteristics of the main studied reservoirs
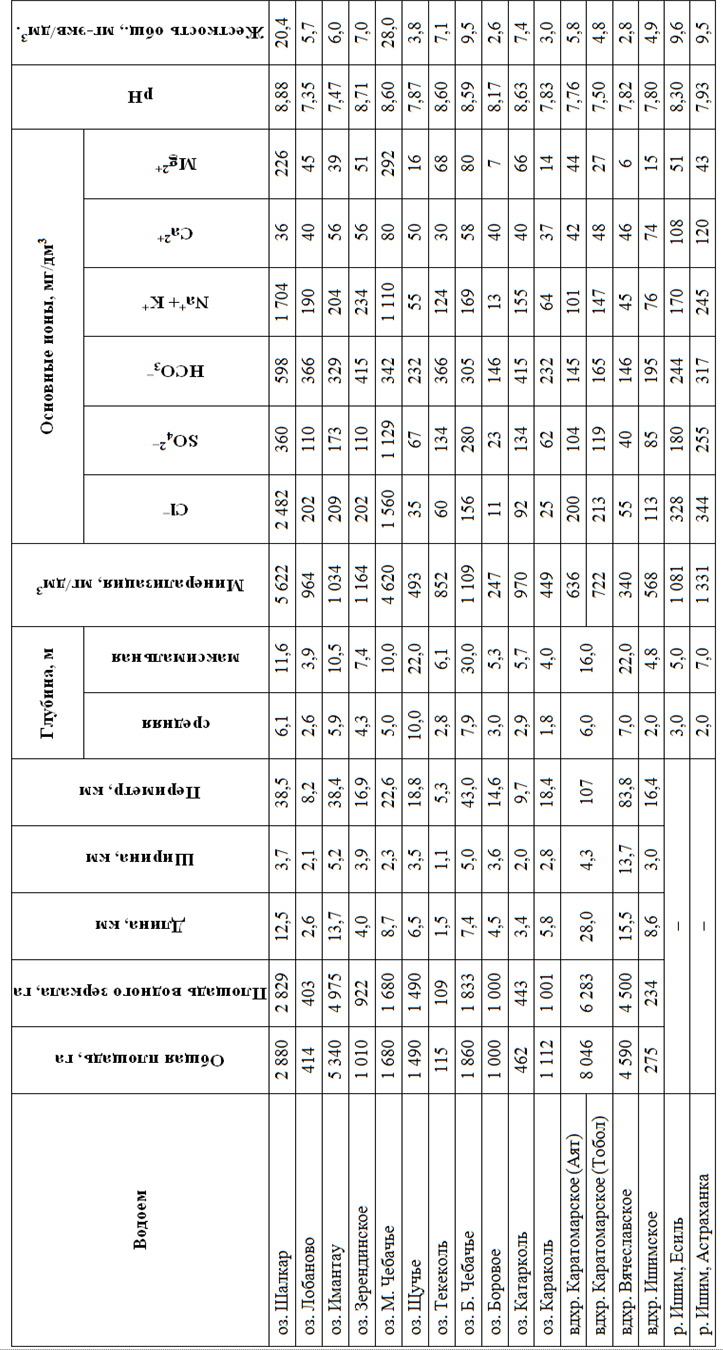
Таблица 2
Table 2
Обратное расчисление линейного роста окуня в водоемах бассейна р. Ишим и сопредельных водных систем
Reverse calculation of the linear growth of perch in the reservoirs of the Ishim River basin and adjacent water systems

Наиболее растянутый возрастной ряд характерен для оз. Лобаново (15+), а также озер Боровое, М. и Б. Чебачье (14+), Шалкар и Катарколь (13+). Для речных и водохранилищных популяций максимальный возраст, отмеченный нами, – одиннадцатый год (10+). Максимально известный для водоемов Казахстана возраст составляет 16 лет [13]. В ареале предельный возраст – до 21 года [14, 15].
Ежегодные натуральные приросты окуня из исследованных водоемов приведены в табл. 3.
Таблица 3
Table 3
Ежегодные натуральные средние приросты длины тела GN, см, окуня из исследованных популяций
Annual natural average increases in body length GN, cm, perch from the studied populations
|
Водоем |
Возраст |
|||||||||||||
|
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
15 |
|
|
оз. Шалкар |
3,5 |
2,8 |
2,6 |
2,4 |
2,2 |
1,8 |
1,7 |
1,6 |
1,5 |
1,8 |
1,2 |
– |
||
|
оз. Лобаново |
3,6 |
3,2 |
2,6 |
2,3 |
2,5 |
2,1 |
2,3 |
1,8 |
1,7 |
1,6 |
1,4 |
1,7 |
||
|
оз. Имантау |
3,3 |
2,7 |
2,4 |
2,0 |
1,8 |
1,6 |
2,7 |
1,6 |
1,1 |
– |
||||
|
оз. Зерендинское |
3,1 |
3,0 |
2,8 |
2,3 |
2,1 |
2,0 |
1,7 |
2,5 |
– |
|||||
|
оз. М. Чебачье |
3,4 |
2,8 |
2,6 |
2,4 |
2,2 |
2,1 |
2,0 |
1,8 |
1,4 |
1,6 |
1,4 |
1,9 |
– |
|
|
оз. Щучье, 2016–2019 г. |
3,3 |
2,9 |
2,5 |
2,1 |
1,7 |
1,6 |
1,3 |
1,4 |
1,3 |
– |
||||
|
оз. Щучье, 2023 г. |
2,8 |
2,9 |
2,7 |
2,2 |
2,0 |
1,9 |
1,5 |
1,3 |
– |
|||||
|
оз. Текеколь |
3,1 |
2,4 |
2,3 |
2,0 |
1,8 |
1,6 |
1,4 |
1,6 |
1,1 |
– |
||||
|
оз. Б. Чебачье |
3,2 |
2,8 |
2,5 |
2,2 |
2,1 |
1,9 |
1,8 |
1,6 |
1,3 |
1,2 |
2,2 |
1,2 |
– |
|
|
оз. Боровое |
3,6 |
2,7 |
2,3 |
2,1 |
2,3 |
2,2 |
2,0 |
2,2 |
1,4 |
2,5 |
1,6 |
– |
||
|
оз. Катарколь |
3,3 |
2,9 |
2,5 |
2,1 |
1,9 |
1,8 |
1,6 |
1,7 |
1,6 |
1,7 |
1,4 |
– |
||
|
оз. Караколь, 2015 г. |
3,2 |
2,7 |
2,5 |
2,1 |
1,9 |
1,7 |
1,9 |
1,6 |
– |
|||||
|
оз. Караколь, 2021 г. |
3,7 |
3,4 |
2,6 |
2,2 |
2,5 |
2,1 |
1,8 |
1,5 |
1,8 |
– |
||||
|
вдхр. Каратомарское (Аят) |
3,3 |
2,8 |
2,2 |
2,1 |
1,5 |
2,3 |
– |
|||||||
|
вдхр. Каратомарское (Тобол) |
3,4 |
3,2 |
2,7 |
2,4 |
2,2 |
1,9 |
1,7 |
1,0 |
– |
|||||
|
вдхр. Вячеславское, 2015 г. |
3,8 |
2,9 |
2,6 |
2,1 |
1,9 |
1,6 |
2,2 |
1,1 |
– |
|||||
|
вдхр. Вячеславское, 2019 г. |
3,6 |
3,2 |
2,5 |
2,0 |
1,8 |
1,7 |
– |
|||||||
|
вдхр. Ишимское, 2014 г. |
3,5 |
2,9 |
2,4 |
2,0 |
– |
|||||||||
|
вдхр. Ишимское, 2021 г. |
3,8 |
3,1 |
3,3 |
2,7 |
2,2 |
1,5 |
1,7 |
– |
||||||
|
р. Ишим, Есиль |
3,9 |
3,0 |
2,2 |
1,8 |
1,9 |
1,3 |
– |
|||||||
|
р. Ишим, Астраханка |
4,0 |
3,1 |
2,5 |
2,2 |
1,9 |
1,5 |
1,9 |
1,2 |
– |
|||||
В целом идет вполне логичное уменьшение натуральных приростов с возрастом. Отмечается интересный факт наибольших приростов на втором году жизни у речных и, отчасти, водохранилищных окуней.
Вместо использования натуральных приростов эффективнее анализировать относительные приросты в ограниченном промежутке лет. В табл. 4 представлены удельные приросты в диапазоне 6 лет.
Таблица 4
Table 4
Удельные средние приросты окуня в диапазоне до 6-летнего возраста
Specific average growth of perch in the range up to 6 years of age
|
Водоем |
Прирост GS6, %, по возрастам |
N, экз. |
|||||
|
1 |
2 |
3 |
4 |
5 |
6 |
||
|
оз. Шалкар |
31,4 |
17,7 |
14,4 |
13,3 |
12,0 |
11,2 |
185 |
|
оз. Лобаново |
27,7 |
17,6 |
15,5 |
14,4 |
12,5 |
12,4 |
40 |
|
оз. Имантау |
31,1 |
16,9 |
14,5 |
13,9 |
12,9 |
10,7 |
35 |
|
оз. Зерендинское |
31,1 |
16,6 |
14,9 |
14,3 |
11,9 |
11,2 |
22 |
|
оз. М. Чебачье |
29,7 |
17,4 |
14,6 |
14,2 |
12,7 |
11,4 |
110 |
|
оз. Щучье, 2016–2019 г. |
31,5 |
17,5 |
14,8 |
14,2 |
12,6 |
9,2 |
54 |
|
оз. Щучье, 2023 г. |
31,8 |
15,3 |
15,7 |
14,5 |
12,0 |
10,7 |
15 |
|
оз. Текеколь |
32,1 |
15,7 |
13,4 |
13,9 |
13,9 |
11,0 |
60 |
|
оз. Б. Чебачье |
32,3 |
16,7 |
14,7 |
13,5 |
11,6 |
11,2 |
240 |
|
оз. Боровое |
31,6 |
17,7 |
15,1 |
12,7 |
11,3 |
11,6 |
27 |
|
оз. Катарколь |
29,6 |
17,2 |
15,8 |
14,4 |
11,8 |
11,3 |
149 |
|
оз. Караколь, 2015 г. |
29,7 |
18,3 |
15,0 |
13,8 |
11,8 |
11,6 |
69 |
|
оз. Караколь, 2021 г. |
29,1 |
19,2 |
16,0 |
12,2 |
11,2 |
12,4 |
13 |
Окончание табл. 4
Ending of Table 4
|
Водоем |
Прирост GS6, %, по возрастам |
N, экз. |
|||||
|
1 |
2 |
3 |
4 |
5 |
6 |
||
|
вдхр. Каратомарское (Аят) |
26,4 |
17,1 |
16,4 |
16,3 |
12,0 |
11,8 |
20 |
|
вдхр. Каратомарское (Тобол) |
25,7 |
18,4 |
16,7 |
14,8 |
12,7 |
11,0 |
15 |
|
вдхр. Вячеславское, 2015 г. |
32,2 |
19,2 |
13,4 |
14,4 |
11,1 |
9,8 |
8 |
|
вдхр. Вячеславское, 2019 г. |
33,1 |
17,9 |
14,3 |
13,7 |
11,2 |
9,7 |
15 |
|
вдхр. Ишимское, 2014 г. |
31,8 |
18,1 |
14,7 |
14,3 |
11,6 |
9,7 |
6 |
|
вдхр. Ишимское, 2021 г. |
27,5 |
18,1 |
14,8 |
16,1 |
13,0 |
10,6 |
7 |
|
р. Ишим, Есиль |
25,6 |
22,2 |
19,5 |
11,1 |
12,0 |
9,6 |
4 |
|
р. Ишим, Астраханка |
30,7 |
19,1 |
16,0 |
13,8 |
10,8 |
9,5 |
41 |
На основании этих данных, используя их как «атрибуты выборки», можно получать различные меры сходства-различия выборок между собой. На основании этих показателей были построены дендрограммы, отражающие близость картины роста в диапазоне 6 лет (рис. 2, 3).
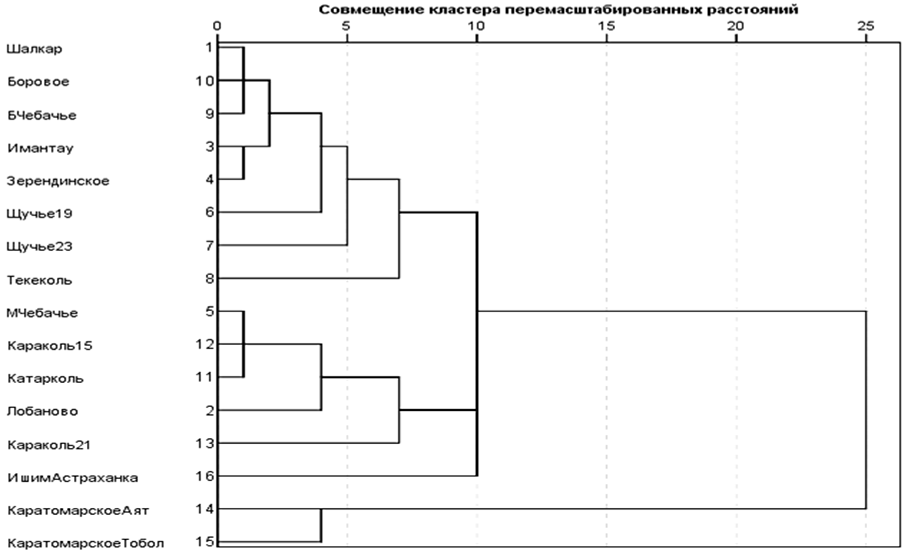
Рис. 2. Дендрограмма с использованием метода межгрупповых связей
сходства выборок по удельным приростам в диапазоне до 6-летнего возраста (GS6)
Fig. 2. Dendrogram using the method of intergroup relations
similarity of samples by specific increment in the range up to 6 years of age (GS6)
Данные части выборок не использовались ввиду малого количества особей. При анализе распределения выборок можно увидеть, что вне зависимости от использованных методов формируются несколько явных кластеров: Шалкар – Боровое – Б. Чебачье; Имантау – Зерендинское; М. Чебачье – Караколь 2015 – Катарколь; Каратомарское (оба) – Лобаново. Также можно объединить обе выборки из оз. Щучье. Прочие группировки не входят в какие-либо объединения. Если сравнить данные табл. 4, можно отметить, что действительно эти объединения формируют некий общий профиль роста на основании очень близких показателей.
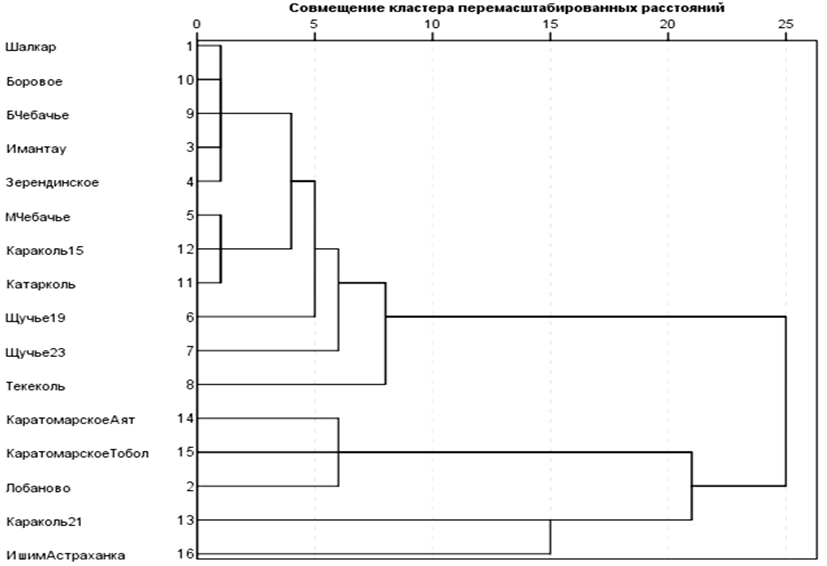
Рис. 3. Дендрограмма с использованием медианного метода
сходства выборок по удельным приростам в диапазоне до 6-летнего возраста (GS6)
Fig. 3. Dendrogram using the median method
of similarity of samples by specific increment in the range up to 6 years of age (GS6)
При построении диаграмм возникает вопрос о хронологических выборках из одного водоема, разделенных временным промежутком более чем одно поколение. Традиционно этот промежуток учитывается как возраст полного становления половозрелости. Как было отмечено выше, для окуня это 3 года. Был проведен анализ изменения роста для 4 популяций (табл. 5): и можно сказать, что для 3 из них динамика не отмечена.
Таблица 5
Table 5
Различия показателей расчисленного роста между хронологическими популяциями (ANOVA)
Differences in calculated growth rates between chronological populations (ANOVA)
|
Водоем |
ANOVA |
Возраст |
|||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
||
|
оз. Щучье |
F |
1,784 |
0,646 |
0,445 |
0,140 |
0,114 |
0,382 |
0,510 |
0,810 |
1,827 |
– |
|
α |
0,184 |
0,423 |
0,506 |
0,709 |
0,736 |
0,536 |
0,482 |
0,385 |
0,264 |
– |
|
|
оз. Караколь |
F |
1,626 |
14,948 |
45,457 |
47,430 |
35,174 |
27,166 |
17,793 |
15,615 |
6,973 |
3,457 |
|
α |
0,205 |
0,0002* |
5 · 10–10 |
4 · 10–10 |
4 · 10–8 |
1 · 10–6 |
7 · 10–5 |
0,0002 |
0,015 |
0,160 |
|
|
вдхр. |
F |
1,137 |
0,005 |
1,218 |
0,080 |
0,005 |
5,453 |
0,980 |
– |
||
|
α |
0,288 |
0,941 |
0,264 |
0,778 |
0,943 |
0,030 |
0,378 |
– |
|||
|
вдхр. |
F |
3,581 |
1,337 |
0,035 |
7,146 |
5,600 |
2,501 |
– |
|||
|
α |
0,063 |
0,564 |
0,852 |
0,010 |
0,028 |
0,142 |
– |
||||
* Жирным шрифтом выделены значения с уровнем достоверности α ≤ 0,001.
Только окуни из оз. Караколь значительно прибавили в скорости роста.
Характер роста окуня из исследованных популяций показывает «преемственность» приростов (табл. 6), т. е. внутри одной выборки расчисленный размер предыдущего года коррелирует с последующим на достаточно высоком уровне.
Таблица 6
Table 6
Коэффициенты корреляции Пирсона индивидуальных показателей расчисленной длины тела между смежными возрастами (α ≤ 0,001)
Pearson correlation coefficients of individual indicators of calculated body length between adjacent ages (α ≤ 0.001)
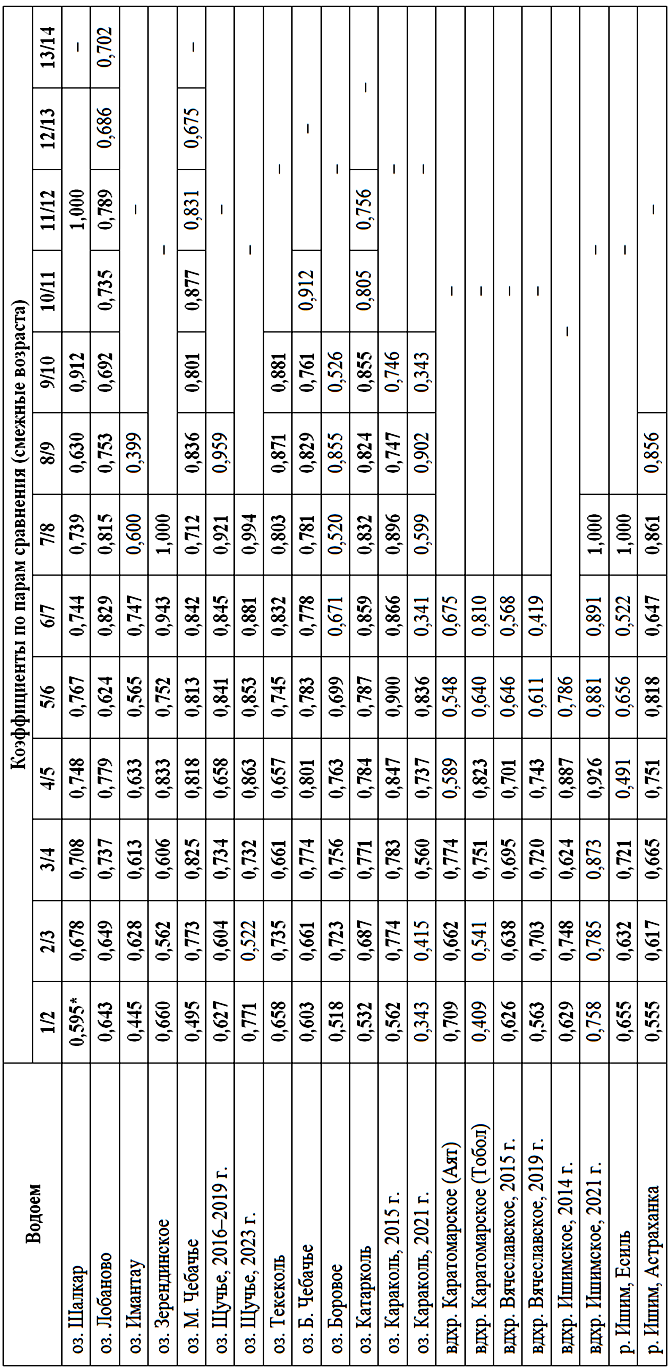
* Жирным шрифтом выделены значения с уровнем достоверности α ≤ 0,001.
Эта корреляция наблюдается почти во всех выборках, за исключением трех. В целом же корреляция расчисленных размеров наиболее ярко просматривается до 5 лет включительно.
Обсуждение
Имеющиеся ретроспективные данные по росту окуня в обозначенном регионе (да и в целом по Казахстану) практически «антикварные». В этой связи возникают существенные проблемы с их использованиием. Так, Л. С. Берг [16] указывает, что видел 8-летнюю икряную самку из оз. Боровое (ныне Акмолинской области, исследованное нами) абсолютной длиной 51 см и весом 2 100 г. То, что он видел ее в июне, не вызывает сомнений, т. к. и у окуня (наряду с некоторыми карповыми) отмечается факт пропуска нереста со зрелыми половыми продуктами
с последующей резорбцией икры. Но вызывает большие сомнения ее возраст, так же, как и вес в 1 950 г в 10+ лет у окуня из оз. Зайсан [17], хотя в этом случае мог сыграть фактор заселения новых мест обитания в образующемся водохранилище.
В этой связи нами не были использованы литературные данные [18] по росту окуня из водоемов исследованного региона. Полученные данные показывают значительную протяженность возрастного ряда у окуней из озер двух национальных парков – «Бурабай» и «Кокшетау». Однако природоохранный статус этих акваторий вряд ли вносит свой вклад
в увеличение продолжительности жизни окуня. Как ни банально, но основную роль здесь играет хорошая изученность и объем собранного материала, что никогда нельзя сбрасывать со счетов.
Для речных группировок вряд ли следует ожидать большей протяженности возрастного ряда, т. к. речная среда обитания более «агрессивная» по отношению к нереофильным видам.
Имеются данные о влиянии площади водоема на рост рыбы [19]. Также давно известно на уровне натуралистических наблюдений, что площадь водоема и сопутствующие факторы (глубина, закоряженность и т. д.) влияют на рост и жизнедеятельность судака, леща и ряда других видов.
Окунь во многом является экологически «средним» видом, не предъявляющим какие-то экстремальные требования к среде обитания и имеющим высокий адаптивный потенциал. Естественно, что будет наблюдаться разница в росте при сравнении водоемов площадью 5 и 500 га. Но в паре 200 и 500 га уже может не быть однозначного ответа. Как и в дан-
ном случае, наиболее протяженный ряд был отмечен для одного из самых малоплощадных водоемов. Вероятно, есть какой-то относительный порог площади водоема, за пределами которого этот фактор не имеет решающего значения.
Что касается величин самого роста у окуня, то они формируются соотношением внутренних группировок с различным темпом роста. В частности, для озер Шалкар и М. Чебачье было описано явление дифференциации внутри группировки по темпам роста [20], которые сразу не бросаются в глаза, как различия между «камышовой» и «пелагической» формами [18, 21]. Соотношения этих «кластеров» регулируется факторами внутренней и внешней природы и имеет серьезное адаптивное значение.
Анализ сходства картин приростов в 6-летнем диапазоне позволяет выделить 2 юнита. Фактически в дискретные юниты группируются выборки из озер нацпарков, кроме Лобаново и вдхр. Каратомарское. К первой группе жестко примыкает выборка из оз. Караколь 2015 г., ко второй – оз. Лобаново, но уровень их связи ниже. Выборки Ишим (Астраханка) и Караколь (2021 г.) не входят в эти 2 объединения, имея собственную картину приростов в этом диапазоне.
Юнит вдхр. Каратомарского выделяют, прежде всего, низкий стартовый рост и повышенные на 3–4 год приросты. По этому набору свойств с ним сближается выборка из оз. Лобаново. У прочих выборок из «озерного» юнита в первый год наблюдаются в основном высокие удельные показатели.
Логически объяснить эти расхождения можно как генетической природой (чувствительностью или предрасположенностью к ответу на какие-либо экологические факторы), так и схожим воздействием экологических факторов. Для сближения выборок из вдхр. Каратомарского и оз. Лобаново, вероятно, причиной будет численность пополнения. Для «озерного» юнита – близкие ответы на воздействие комплекса экологических факторов, что уже предполагает эпигенетический ответ.
В целом данный вопрос очень интересен. Его решение содержит в себе большую практическую составляющую. Однако пока ему не уделялось должного значения даже на уровне решения логических задач.
Сравнение временных выборок из одного водоема показывает достаточную консервативность роста. Для вдхр. Вячеславского ранее [22] на основании эмпирических подходов было высказано мнение об отсутствии различий в росте у окуня из хронологических выборок. Эта консервативность может основываться только на единообразии экологических факторов на протяжении определенного периода. Если эти факторы меняются, происходит изменение картины роста, что и произошло на оз. Караколь. Повышение водности вызвало увеличение площадей кормовых стаций и, как следствие, повышение темпов роста. Следует обратить внимание, что рост в первый год жизни не обладает достоверными отличиями. В этом случае, вероятно, показатели роста зависят от численности этой генерации, т. е. от процессов воспроизводства. Примерно одинаковая величина пополнения и питание зоопланктоном [23–26], обладающим относительно стабильной для потребления окунем частью биомассы, формируют примерно одинаковый рост. Далее во многих водоемах окунь переходит на потребление бентоса и нектона. Это может произойти уже на первый год жизни [27–31]. Для оз. Караколь специальных исследований на эту тему не проводилось, но нет оснований предполагать, что здесь это происходит как-то иначе. Новый трофический переход формирует более жесткую конкурентную среду как за счет большой исходной численности генерации, так и за счет более активного избегания хищников жертвами. И далее, при переходе на хищничество, конкуренция продолжается до момента снятия одного из условий – высокой численности генераций. А рост позволяет расширить кормовую базу за счет введения в оборот новых размерных классов жертв, соответственно, темпы роста сглаживаются. Это не означает, что все достигают примерно равного размера, но приросты становятся примерно равными, насколько позволяет индивидуальная природа особи.
Некоторые тенденции к дифференциации хронологических выборок имеются и для вдхр. Ишимского. Выборки 2021 г. из этих 2-х водоемов (оз. Караколь и вдхр. Ишимское) не проявляют взаимосвязи между смежными годами. Из-за резкого улучшения гидрологического режима в середине 2010-х гг. исчезла канализация роста за счет дефицита трофических ресурсов, поэтому рост стал зависеть от индивидуальных способностей организма. Безусловно, подобное могло произойти на фоне невысокой экологически эффективной численности вида в водоеме, не обеспечивающей занятие всех пространственных ниш.
Корреляции между смежными возрастами в других выборках достаточно существенны, что показывает определенный уровень стабильности воздействий на процессы роста.
Рост окуня из водоемов Ишимского бассейна эмпирически отличается меньшими темпами от ранее описанных нуринских выборок [4]. Вероятнее всего, это связано с типом изученных популяций в бассейнах. В случае Нура-Сарысуйского бассейна исследовались осваиваемые промысловым ловом, в случае Ишимского – более развито любительское (рекреационное) рыболовство. В первом случае генерационный ряд короче, а темпы роста выше, во втором – ряд поколений более растянут, темпы роста ниже.
Заключение
Таким образом, рост окуня в казахстанской части бассейна р. Ишим несколько уступает таковому у окуня из бассейна р. Нуры. Вместе с тем возрастной ряд более протяженный именно у ишимских выборок. Это, в принципе, объясняется тем, что в последнем случае были исследованы в основном промыслово-эксплуатируемые популяции, а в ишимской системе – рекреационные водоемы с доминированием любительского лова.
Не было выявлено каких-либо существенных доказательств влияния гидроморфологических показателей водоемов на рост окуня. Но в процессе работы было показано изменение темпов роста в результате изменения гидрологических показателей. В новой, нестабильной ситуации у окуня произошло не только увеличение темпов роста, но
и изменилась его регуляция – она стала менее строгой. В стабильных условиях, наблюдаемых у большинства других выборок, рост достаточно «канализирован» и наблюдается преемственность между генерациями.
Анализ сходства-различия выборок показал наличие двух типов распределения темпов роста внутри жизненного цикла. Относительным эталоном приходится признать картину роста группы, куда включены практически все озерные популяции. Здесь идет плавное уменьшение удельного роста с возрастом. Такое же уменьшение характерно и для второй группы, но в первый год удельный рост минимальный, по сравнению с первой группой, а в 3-4 годы жизни эти показатели становятся практически максимальными (по сравнению с ней же). Это явление, вероятно, формируется как экологическими, так и эпигенетическими факторами.
1. Казахстан. Национальная энциклопедия. Алматы: Қазақ эниклопедиясы, 2005. Т. 2. 560 с.
2. Митрофанов В. П., Дукравец Г. М., Песериди Н. Е., Полторыхина А. Н., Ерещенко В. И., Захаров С. С., Мельников В. А., Солонинова Л. Н., Орлова И. В., Горюнова А. И. Рыбы Казахстана. Алма-Ата: Наука, 1986. Т. 1. 272 с.
3. Le Cren E. D. The Determination of the Age and Growth of the Perch (Perca fluviatilis) from the Opercular Bone // Journal of Animal Ecology. 1947. V. 16. N. 2. P. 188–204.
4. Крайнюк В. Н., Асылбекова С. Ж., Шуткараев А. В. Линейный рост окуня Perca fluviatilis L., 1758 (Percidae) в бассейнах рек Нура и Сарысу // Вестн. Астрахан. гос. техн. ун-та. Сер.: Рыбное хозяйство. 2020. № 3. С. 83–95. DOI:https://doi.org/10.24143/2073-5529-2020-3-83-95.
5. Francis R. I. C. C. Back-calculation of fish length: a critical review // J. Fish. Biol. 1990. V. 36. N. 6. P. 883–902.
6. Мина М. В., Клевезаль Г. А. Рост животных. М.: Наука, 1976. 291 с.
7. Ильмаст Н. В. Введение в ихтиологию. Петрозаводск: Изд-во КарНЦ РАН, 2005. 148 с.
8. Дюран Ю., Оделл П. Кластерный анализ. М.: Статистика, 1977. 128 с.
9. Мандель И. Д. Кластерный анализ. М.: Финансы и статистика, 1988. 176 с.
10. Плохинский Н. А. Биометрия. М.: Изд-во МГУ, 1970. 367 с.
11. Коросов А. В., Горбач В. В. Компьютерная обработка биологических данных. Петрозаводск: Изд-во ПГУ, 2007. 76 с.
12. Бююль А., Цёфель П. SSPS: Искусство обработки информации. СПб.: ДиаСофтЮП, 2005. 608 с.
13. Горюнова А. И. Формирование ихтиофауны Джезказганского водохранилища // Сб. работ по ихтиологии и гидробиологии. Алма-Ата: Изд-во АН КазССР, 1956. Вып. 1. С. 31–73.
14. Craig J. F. Percids fishes. Systematics, Ecology and Exploitation. Oxford: Blackwell Science Ltd, 2000. 352 p.
15. Kottelat M., Freyhof J. Handbook pf European Freshwater Fish. Berlin: Publications Kottelat, Cornol and Freyhof, 2007. 646 p.
16. Берг Л. С. Рыбы пресных вод СССР и сопредельных стран. М.-Л.: Изд-во АН СССР, 1949. Т. 3. С. 927–1382.
17. Мартехов П. Ф. Биологические основы создания стада ценных промысловых рыб на Зайсане в связи с образованием Бухтармино-Зайсанского водохранилища // Биологические основы рыбного хозяйства: тр. Всесоюз. совещ. Томск, 1959. С. 191–200.
18. Митрофанов В. П., Дукравец Г. М. и др. Рыбы Казахстана. Алма-Ата: Наука, 1989. Т. 4. 312 с.
19. Durham B. W., Pope K. L., Wilde G. R. Factors Affecting Regional Variation in Growth of Channel Catfish // North American Journal of Fisheries Management. 2005. V. 25. N. 1. P. 93–97. DOI:https://doi.org/10.1577/M03-129.1.
20. Krainyuk V., Shutkarayev A., Isbekov K., Assylbekova S., Kulikov Ye. Intra-group growth variability of perch (Perca fluviatilis L., 1758) (Percidae) of Shalkar and Small Chebachye lakes // AACL Bioflux. 2023. V. 16. N. 5. P. 2757–2764.
21. Никольский Г. В. Рыбы Аральского моря. М., 1940. Вып. 1 (XVI). 216 с.
22. Крайнюк В. Н. Внутригрупповая изменчивость роста окуня Perca fluviatilis L., 1758 (Percidae) водохранилища Вячеславского // Рыбоводство и рыбное хозяйство. 2021. № 11. С. 8–15. DOI:https://doi.org/10.33920/sel-09-2111-01.
23. Глушакова В. И. О питании молоди основных промысловых видов рыб Бухтарминского водохранилища на ранних этапах развития // Биологические основы рыбного хозяйства республик Средней Азии и Казахстана. Фрунзе: Илим, 1981. С. 252–254.
24. Козляткин А. Л. Значение акклиматизированных ракообразных в питании окуня и молоди судака Бухтарминского водохранилища // Биологические основы рыбного хозяйства республик Средней Азии и Казахстана. Ашхабад: Ылым, 1974. С. 66–67.
25. Kratochvíl M., Peterka J., Kubečka J., Matěna J., Vašek M., Vaničková I., Čech M., Seďa J. Diet of larvae and juvenile perch, Perca fluviatilis performing diel vertical migrations in a deep reservoir // Folia Zoologica. 2008. V. 57. N. 3. P. 313–323.
26. Skrzypczak A., Mamcarz A., Kujawa R., Ku-charczyk D., Furgala-Selezniow G. Feeding habits of larval Eurasian perch, Perca fluviatilis (Percidae) // Italian Journal of Zoology. 1998. V. 65. Supl. 1. P. 243–245. DOI:https://doi.org/10.1080/11250009809386825.
27. Горлачева Е. П. Питание рыб озера Кенон // Природоохранное сотрудничество в приграничных экологических регионах: Россия – Китай – Монголия. Чита: Поиск, 2012. Ч. 1. С. 114–119.
28. Тютеньков С. К. Питание и пищевые взаимоот-ношения рыб оз. Кургальджин // Сб. работ по ихтиоло-гии и гидробиологии. Алма-Ата: Изд-во АН КазССР, 1956. Вып. 1. С. 155–171.
29. Adámek Z., Musil J., Sukop I. Diet composition and selectivity in O+ perch (Perca fluviatilis L.) and its competition with adult fish and carp (Cyprinus carpio L.) stock in pond culture // Agriculturae Conspectus Scientificus. 2004. V. 69. N. 1. P. 21–27.
30. Amundsen P.-A., Bøhn T., Popova O. A., Staldvik F. J., Reshetnikov Yu. S., Kashulin N. A., Lukin A. A. Ontogenetic niche shifts and resource partitioning in a subarctic piscivore fish guild // Hydrobiologia. 2003. V. 497. N. 1. P. 109–119. DOI:https://doi.org/10.1023/A:1025465705717.
31. Lappalainen A., Rask M., Koponen H., Vesala S. Relative abundance, diet and growth of perch (Perca fluvi-atilis) and roach (Rutilus rutilus) at Tvaerminne, northern Baltic Sea, in 1975 and 1997: responses to eutrophication? // Boreal Environ. Research. 2001. V. 6. N. 2. P. 107–118.
