











Russian Federation
Russian Federation
Russian Federation
Russian Federation
The growing conditions of sturgeon fish affect the growth and physiological state, their morphological characteristics. The study of changes in physiological processes under the influence of a complex of factors in the conditions of cage complexes and closed water supply systems will allow for timely adjustment of technology and reduce risks when growing commercial products. The repair and breeding herds of fish breeding enterprises contain sterlet individuals of different ages, which annually, upon reaching puberty, enter the fish breeding process as producers. In this regard, we have studied the main morphophysiological indicators of two-year-olds kept in cages and in closed water supply installations (CWSI). The influence of environmental factors was noted not only on growth indicators, but also on the size of the internal organs of fish and their condition. The unstable temperature regime in the cages, dependence on unpredictable changes in the hydrochemical parameters of the aquatic environment contributes to deep physiological transformations of an adaptive nature, which slows down the growth rate and accumulation of reserve substances. Changes in morphometric parameters were also evident. In sturgeon, there is an increase in the relative weight of the heart, spleen and gonads, a decrease in the weight of the kidneys and liver. The influence of optimal temperature and trophic conditions on the index of the gastrointestinal tract was noted, which was slightly lower in fish from the CWSI than in individuals exposed to negative factors in cages. Sterlet blood counts, regardless of the conditions of detention, corresponded to physiologically healthy individuals. The creation of optimal hydrological and trophic conditions in CWSI contribute to the accumulation of phospholipids and linolenic fatty acids in tissues, which play an important role in the adaptation of the body to negative environmental factors.
sterlet, physiological state, growing conditions, morphophysiological parameters, indices of internal organs, blood, lipids, fatty acids
Введение
Одним из наиболее распространенных объектов выращивания в товарном осетроводстве является стерлядь, обладающая высокой степенью адаптивности к внешним условиям, более раним, в отличие от других видов осетровых, созреванием, небольшим интервалом между нерестами, высоким темпом роста, ценными потребительскими свойствами. Выращивание данного вида осетровых рыб проводят как в условиях садковых хозяйств, так и в установках замкнутого водоснабжения (УЗВ), а также в прудах и небольших водоемах. Условия выращивания оказывают значительное влияние на рост и состояние рыб, их морфологические признаки.
В настоящее время интенсивность загрязнения промышленными и сельскохозяйственными стоками естественной среды обитания рыб постоянно возрастает [1], что ведет к регрессу сложившихся биоценозов и, зачастую, к необратимым изменениям физиологического состояния особей. Постоянное присутствие загрязнителей в водной среде препятствует нормальному росту и развитию организма рыб, способствует возникновению аномалий и нарушению обменных процессов. Подобное влияние на организм рыб оказывают и факторы стресса, которые являются неотъемлемой частью рыбоводного процесса [2]. Таким образом, в товарном осетроводстве в условиях садковых предприятий на состояние выращиваемых объектов негативное воздействие оказывает комплекс факторов антропогенной природы, которые способствуют нарушению гомеостаза внутренней среды организма рыб, проявляющемуся
в изменении морфофизиологических показателей и снижении их продуктивных качеств.
Высокая способность стерляди адаптироваться к изменяющимся факторам внешней среды позволяет компенсировать степень воздействия и предотвратить критические изменения физиологического состояния. Адаптационный потенциал организма зависит от особенности метаболизма в тот или иной период жизненного цикла [2]. Этот факт свидетельствует о регулярной трансформации обменных процессов в организме рыб не только в течение жизни, но и в результате постоянного воздействия внешних факторов. В раннем периоде постэмбриогенеза обмен веществ в организме направлен на накопление и использование пищевых нутриентов на рост и развитие, в период созревания энергетический потенциал направлен на формирование гонад [3, 4]. Изучение изменений физиологических процессов при воздействии комплекса факторов в условиях садковых комплексов и систем замкнутого водоснабжения позволит проводить своевременную корректировку технологии выращивания и снижать риски при получении товарной продукции.
Целью исследований является изучение морфофизиологических показателей стерляди, выращиваемых в условиях садковых хозяйств и при регулируемых условиях водной среды УЗВ.
Материал и методы исследований
Основой представленных материалов являются обобщенные многолетние данные, собранные на садковых хозяйствах Нижней Волги и в УЗВ (уникальной научной установке (УНУ) НТИ РФ Рег. № 3662433) «Научно-исследовательский комплекс передовых технологий аквакультуры и гидроэкологии» факультета биотехнологий и рыбного хозяйства Московского государственного университета технологий и управления имени К. Г. Разумовского (ПКУ).
Объектом исследования являлись двухлетки стерляди, содержащиеся в садках и в условиях УЗВ, средней массой 318,1 ± 15 и 335,5 ± 25 г соответственно. Для исследований отбиралось по 20 особей без видимых повреждений.
Оценку физиологического состояния стерляди проводили по основным гематологическим и физиолого-биохимическим показателям. Отбор крови у рыб осуществляли из хвостовой артерии прижизненным способом. Содержание гемоглобина в крови измеряли фотометрическим методом. Среднюю концентрацию гемоглобина в эритроците рассчитывали по И. И. Гительзону и И. А. Терскову [5], подсчет эритроцитов – в камере Горяева, скорость оседания эритроцитов – в аппарате Панченкова. Определение клеток осуществляли по классификации Н. Т. Ивановой [6]. Общий сывороточный белок определяли рефрактометрически.
Химический состав тканей рыб анализировали стандартными методами [7]. Для извлечения общих липидов применяли модифицированный метод, основанный на технике Блайя и Дайера. Липиды далее разделяли на классы с использованием тонкослойной хроматографии, а их разгонку осуществляли в системе растворителей [8]. Фосфолипиды разделяли на колонках с модифицированными целлюлозами. Концентрацию холестерина определяли методом Либермана – Бурхарда, а эфиры стеринов – по реакции с хромотроповой кислотой, следуя методике, указанной в работе Н. А. Абросимовой и соавторов [9].
Результаты исследований
В ремонтно-маточных стадах рыбоводных предприятий содержатся разновозрастные особи стерляди, которые ежегодно при достижении половой зрелости входят в рыбоводный процесс в качестве производителей. В связи с этим нами исследованы основные морфофизиологические показатели двухлеток, содержащихся в садках и в УЗВ. Содержание рыб в садках происходит при естественном температурном режиме, в условиях, способствующих реализации природного потенциала роста и развития. В УЗВ в условиях оптимального термического режима и сбалансированного кормления интенсивность обменных процессов повышается, что способствует увеличению интенсивности роста рыб [10], сокращению межнерестового интервала [11]. Влияние факторов среды обитания отмечено не только на ростовые показатели, но и на размеры внутренних органов рыб и их состояние.
Исследованные двухлетки стерляди, содержащиеся в условиях садковых хозяйств, по степени накопления питательных веществ были менее упитанными. Коэффициент упитанности этих рыб был в среднем на 14 % ниже, чем у особей, выращенных и содержащихся в условиях УЗВ. Нестабильный температурный режим в садках, зависимость от непрогнозируемых изменений гидрохимических показателей водной среды способствуют глубоким физиологическим преобразованиям адаптивного характера, что замедляет темп роста и кумуляцию резервных веществ.
Очевидными были и изменения морфометрических показателей. Индекс печени рыб, выращенных при нерегулируемом гидрологическом режиме, был ниже на 12 % и соответствовал в среднем 2,23 ± 0,41 %. При замкнутом цикле водообеспечения в условиях с регулируемыми основными показателями качества водной среды гепатосоматический индекс в среднем составлял 2,87 ± 0,15 %. Противоположная закономерность выявлена при изучении кардиосоматических индексов. Наибольший индекс сердца отмечен у рыб в садковых хозяйствах. Его величина была выше на 20 %, чем у рыб из установки с замкнутым циклом водообеспечения. Полученные результаты согласуются с ранее опубликованными сведениями Т. В. Бехтеревой [12], Д. Ф. Нгуен [13], которые утверждают, что стрессовые факторы оказывают влияние на морфометрические показатели, проявляющееся в повышении относительной массы сердца, селезенки и гонад, снижении массы почек и печени. Морфофизиологические показатели стерляди, содержащейся в УЗВ и садках, представлены на рис. 1.

Рис. 1. Морфофизиологические показатели стерляди, содержащейся в УЗВ и садках
Fig. 1. Morpho-physiological parameters of sterlet kept in CWSI and cages
Отмечено влияние оптимальных температурного и трофического режимов на индекс желудочно-кишечного тракта. У рыб из УЗВ индекс ЖКТ был несколько ниже, чем у особей, испытывающих воздействие негативных факторов в садках. Данное обстоятельство объясняется опережающим увеличением длины тела относительно роста кишечника, что обусловлено высокой эффективностью усваивания питательных веществ корма и метаболизма рыбы.
Таким образом, проведенные исследования доказывают влияние неблагоприятных факторов внешней среды в условиях садкового выращивания, главным образом температуры воды, на морфометрические показатели внутренних органов стерляди.
Воздействие на рыб внешних факторов среды приводит к изменению скорости и характера обменных процессов, переход которых за пределы адаптационной способности организма приводит к патологическим нарушениям и, как следствие, к гибели рыб [14, 15]. Неблагоприятные условия среды обитания рыб, вызывающие стресс, сопровождаются изменениями состава крови, которые наиболее точно отражают физиологическое состояние и наличие патологических изменений. Комплексное исследование гематологических показателей рыб позволяет своевременно выявить дисбаланс в организме и правильно диагностировать заболевание [16].
Показатели красной крови стерляди из садковых хозяйств находились в пределах физиологической нормы здоровых рыб. Содержание гемоглобина в крови колебалось в пределах от 77,1 ± 0,7 до 81,4 ± ± 0,5 г/л, количество эритроцитов – от 0,902 · 106 до 1,02 · 106 кл/мкл. Реакция оседания эритроцитов свидетельствовала об отсутствии воспалительных процессов в организме рыб и соответствовала нормальным значениям (2,6–3,4 мм/ч). Показатели крови стерляди из УЗВ соответствовали показателям физиологически здоровых особей [17].
Форменный состав элементов периферической крови стерляди в УЗВ и при садковом выращивании представлен на рис. 2 и в табл. 1.
а б
Рис. 2. Картина крови особей стерляди, выращиваемых в садках (а) и УЗВ (б)
Fig. 2. Blood sample of sterlet individuals cultured in cages (a) and CWSI (б)
Таблица 1
Table 1
Гематологические показатели ремонтной группы стерляди, %
Hematological parameters of the sterlet repair group, %
|
Показатель |
Группа |
|
|
УЗВ |
Садки |
|
|
Лимфоциты |
86,1 ± 1,3 |
85,65 ± 1,6 |
|
Моноциты |
3,42 ± 0,8 |
3,3 ± 0,5 |
|
Нейтрофилы |
7,8 ± 0,30 |
7,9 ± 0,66 |
|
Эозинофилы |
3,1 ± 0,55 |
3,5 ± 0,3 |
|
Тромбоциты |
1,9 ± 0,3 |
2,1 ± 0,4 |
Существенной разницы по относительному количеству элементов белой крови у стерляди при разных режимах выращивания выявлено не было. При этом количество эозинофилов у рыб в условиях садкового содержания было несколько выше, что может быть обусловлено стимуляцией иммунного ответа на неблагоприятные факторы водной среды.
Количество сывороточного белка зависело от условий содержания рыб. Наличие факторов стресса при садковом выращивании способствовало снижению этого показателя до 27,3 г/л. Существенное снижение показателя общего сывороточного белка в крови приводит к снижению жизнестойкости рыб и патологическим нарушениям физиологических процессов. Количество белка в крови рыб, содержащихся в оптимальных условиях УЗВ, колебалось в незначительных пределах и составляло в среднем от 30,1 до 35,3 г/л, что свидетельствует о нормальном состоянии здоровья выращиваемых рыб при благоприятных режимах содержания и кормления.
Биохимический состав тела стерляди из садкового хозяйства отличался большим количеством белка в сухом веществе и низким уровнем липидов. По показателям биохимического состава тела состояние рыб как в садках, так и в УЗВ оценивается удовлетворительным (рис. 3).

Рис. 3. Биохимический состав тканей ремонтной группы стерляди
Fig. 3. Biochemical composition of tissues of sterlet breeding group
Содержание фосфолипидов в общих липидах тканей стерляди, содержащейся в УЗВ и садках, составляло 41,3 и 39,8 % соответственно. На рис. 4 представлены результаты исследования состава общих липидов тела двухлеток стерляди.
В липидах тканей рыб, выращенных в УЗВ, наблюдалось повышение концентрации жирных кислот линоленового ряда на 7 %, что обусловлено условиями содержания (табл. 2).
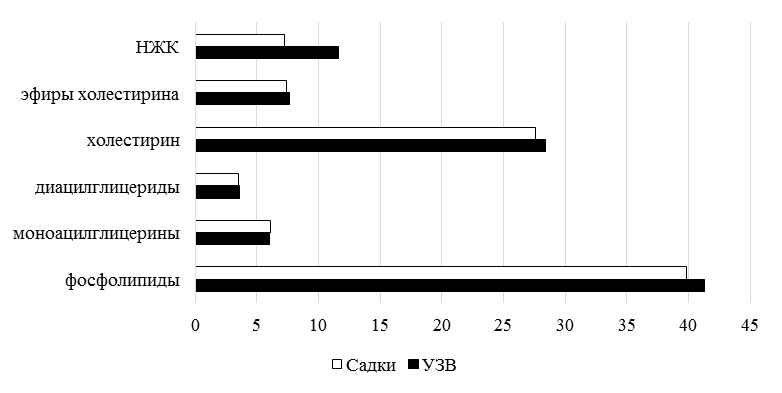
Рис. 4. Общие липиды тканей ремонтной группы стерляди
Fig. 4. Common lipids of sterlet repair group tissues
Таблица 2
Table 2
Состав жирных кислот стерляди, выращиваемой в УЗВ и в садковых хозяйствах, %
The composition of fatty acids of sterlet grown in CWSI and in the conditions of cage farms, %
|
Условия |
Жирные кислоты |
|||||
|
Насыщенные |
Ненасыщенные |
|||||
|
Моноеновые |
Полиеновые |
Σ ω3 |
Σ ω6 |
ω3 / ω6 |
||
|
УЗВ |
34,5 ± 1,8* |
32,1 ± 1,0 |
33,4 ± 1,2 |
17,4 ± 0,3* |
16,0 ± 0,4 |
1,1 |
|
Садки |
32,4 ± 1,2* |
33,4 ± 0,8 |
34,2 ± 1,1 |
18,5 ± 0,6* |
15,7 ± 0,5 |
1,2 |
* Достоверность различий p < 0,05.
Высокая степень обеспеченности фосфолипидов полиненасыщенными жирными кислотами линоленового ряда стимулирует защитные механизмы, увеличивая устойчивость организма к различным заболеваниям и обеспечивая реализацию адаптивных процессов к стрессовым факторам [18].
Полученные гематологические и биохимические показатели соответствуют ранее опубликованным результатам исследований Г. Ф. Металлова, В. М. Распопова и др. [19], А. А. Корчунова и др. [11], А. Б. Ахмеджановой с соавторами [16] и др.
Заключение
Определена зависимость состава жирных кислот в тканях стерляди от условий выращивания. Создание оптимальных гидрологических и трофических условий в установках замкнутого водоснабжения способствует накоплению в тканях фосфолипидов и жирных кислот линоленового ряда, играющих важную роль в адаптации организма к негативным факторам внешней среды. В целом физиологическое состояние исследуемых особей стерляди, содержащихся в ремонтных стадах при различных гидрологических условиях, было удовлетворительным.
1. Amanov Sh. M., Evdokimov E. V. Sostoianie osetrovykh ryb v Kaspiiskom more [The condition of stur-geon fish in the Caspian Sea]. Universum: khimiia i biologiia, 2022, no. 12-1 (102), pp. 15-18.
2. Shul'man G. E. Fiziologo-biokhimicheskie osobennosti godovykh tsiklov ryb [Physiological and biochemical features of annual fish cycles]. Moscow, Pishchevaia promyshlennost' Publ., 1972. 367 p.
3. Shatunovskii M. I. Ekologicheskie zakonomernosti obmena veshchestv morskikh ryb [Ecological patterns of marine fish metabolism]. Moscow, Nauka Publ., 1980. 283 p.
4. Metallov G. F., Ponomareva E. N., Sorokina M. N., Grigor'ev V. A., Korchunov A. A. Funktsional'naia napravlennost' biokhimicheskikh protsessov u samok gibrida sterliad' × beluga (Acipenser ruthenus Linnaeus, 1758 × Huso huso Linnaeus, 1758) v reproduktivnom tsikle [Functional orientation of biochemical processes in females of the sterlet × beluga hybrid (Acipenser ruthenus Linnaeus, 1758 × Huso huso Linnaeus, 1758) in the reproductive cycle]. Doklady Akademii nauk, 2018, vol. 478, no. 6, pp. 727-729.
5. Gitel'zon I. I., Terskov I. A. O sposobe vyrazheniia gemoglobina v eritrotsite [About the method of expression of hemoglobin in erythrocyte]. Laboratornoe delo, 1956, no. 6, pp. 6-10.
6. Ivanova N. T. Atlas kletok krovi ryb [Atlas of fish blood cells]. Moscow, Pishchevaia promyshlennost' Publ., 1983. 80 p.
7. Shcherbina M. A. Metodicheskie ukazaniia po fiziologicheskoi otsenke pitatel'noi tsennosti kormov dlia ryb [Guidelines for the physiological assessment of the nutritional value of fish feed]. Moscow, Izd-vo VNIIPRKh, 1983. 83 p.
8. Alekseev S. M., Tarablin M. B., Sarycheva I. K., Evstigneeva R. P. Gazokhromatograficheskii analiz metilo-vykh efirov zhirnykh kislot na kolonkakh so smeshannym sorbentom [Gas chromatographic analysis of methyl esters of fatty acids on columns with a mixed sorbent]. Khimiko-farmatsevticheskii zhurnal, 1981, vol. 15, pp. 118-122.
9. Abrosimova N. A., Abrosimov S. S., Saenko E. N. Kormovoe syr'e i dobavki dlia ob"ektov akvakul'tury [Feed raw materials and additives for aquaculture facilities]. Rostov-on-Don, Everest Publ., 2005. 144 p.
10. Khrustalev E. I., Molchanova K. A., Abdulrakhman Z. Kh. Osobennosti raskrytiia rostovoi potentsii u raznykh ob"ektov akvakul'tury [Features of the disclosure of growth potency in different aquaculture facilities]. Vodnye bioresursy, akvakul'tura i ekologiia vodoemov: trudy V Baltiiskogo morskogo foruma. Vserossiiskaia nauchnaia konferentsiia (Kaliningrad, 23–24 maia 2017 g.). Kaliningrad, Izd-vo KGTU, 2017. Pp. 154-158.
11. Korchunov A. A., Metallov G. F., Grigor'ev V. A., Kovaleva A. V. Dinamika biokhimicheskogo sostava tela i polovykh produktov sterliadi (Acipenser ruthenus Linnaeus, 1758) estestvennykh populiatsii i vyrashchennykh v ustanovkakh zamknutogo vodosnabzheniia [Dynamics of the biochemical composition of the body and sexual products of sterlet (Acipenser ruthenus Linnaeus, 1758) of natural populations and grown in closed water supply installations]. Vestnik Astrakhanskogo gosudarstvennogo tekhnicheskogo universiteta. Seriia: Rybnoe khoziaistvo, 2012, no. 1, pp. 136-143.
12. Bekhtereva T. V. Ekologo-fiziologicheskie issledo-vaniia [Ecological and physiological research]. Moscow, Nauka Publ., 1998. 121 p.
13. Nguen D. F., Kurmanbaev A. B. Kachestvennyi sostav osetra, migriruiushchego v r. Volgu v 2005 g. [The qualitative composition of the sturgeon migrating to the Volga River in 2005]. Tezisy dokladov IX Ezhegodnoi nauchnoi konferentsii studentov i aspirantov bazovykh kafedr Iuzhnogo nauchnogo tsentra RAN (Rostov-na-Donu, 11–24 aprelia 2013 g.). Rosov-on-Don, 2013. P. 46.
14. Moiseenko T. I. Izmenenie strategii zhiznennogo tsikla ryb pod vozdeistviem khronicheskogo zagriazneniia vod [Changing the life cycle strategy of fish under the influence of chronic water pollution]. Ekologiia, 2002, no. 1, pp. 50-60.
15. Faizulina D. R., Bazeliuk N. N., Aksenov V. P. Nekotorye aspekty patologichnykh znachenii biokhimich-eskikh pokazatelei krovi vobly Rutilus rutilus caspicus i leshcha Abramis brama v sovremennykh ekologicheskikh usloviiakh Volzhsko-Kaspiiskogo basseina [Some aspects of pathological values of biochemical blood parameters of roach Rutilus rutilus caspicus and bream Abramis brama in modern ecological conditions of the Volga-Caspian basin]. Trudy VNIRO, 2016, vol. 162, pp. 20-27.
16. Akhmedzhanova A. B., Ponomarev S. V., Viatchin V. V., Fedorovykh Iu. V., Levina O. A., Dutikov E. A. Otsenka bioindikatorov fiziologo-biokhimicheskikh poka-zatelei remontno-matochnogo stada osetrovykh ryb [Evaluation of bioindicators of physiological and biochemical parameters of the repair and brood stock of sturgeon fish]. Rybnoe khoziaistvo, 2021, no. 5, pp. 97-100. DOI:https://doi.org/10.37663/0131-6184-2021-5-97-100.
17. Asadi F., Masoudifard M., Vajhi A., Lee K., Pourkabir M., Khazraeiniaet P. Serum biochemical parameters of Acipenser persicus. Fish Physiology and Biochemistry, 2006, vol. 32, pp. 43-47.
18. Fokina N. N., Lysenko L. A., Ruokolainen T. R., Sukhovskaia I. V. Zavisimost' soderzhaniia lipidov i nena-syshchennykh zhirnykh kislot v skeletnykh myshtsakh raduzhnoi foreli ot uslovii vyrashchivaniia i fiziologicheskogo sostoianiia ryb [The dependence of the content of lipids and unsaturated fatty acids in the skeletal muscles of rainbow trout on the growing conditions and the physiological state of the fish]. Prikladnaia biokhimiia i mikrobiologiia, 2020, vol. 56, no. 3, pp. 305-312.
19. Metallov G. F., Raspopov V. M., Aksenov V. P., Chipinov V. G. Biokhimicheskie i morfofiziologicheskie pokazateli russkogo osetra v sovremennykh ekologicheskikh usloviiakh Volgo-Kaspiia [Biochemical and morphophysiological parameters of the Russian sturgeon in modern environmental conditions of the Volga-Caspian Sea]. Teplovodnaia akvakul'tura i biologicheskaia produktivnost' vodoemov aridnogo klimata: sbornik materialov i dokladov Mezhdunarodnogo simpoziuma. Astrakhan', 2007. Pp. 484-486.
