











Russian Federation
Russian Federation
The article highlights physiological and biochemical parameters of atherina (Atherina boyeri caspia Risso, 1810) in the Middle Caspian Sea at various stages of the annual cycle. The state of the fish organism was studied in the period from February to October 2022, which characterized the pre-spawning, spawning and feeding periods. The size-weight and age parameters of atherina were determined. The concentration of the main reserve substances of the fish body - total lipids and water-soluble protein – was analyzed in the muscles of the atherina. In the pre-spawning period and at the beginning of spawning of females, the level of morphological changes in the gonads was studied. There were no found significant differences in the content of total lipids and water-soluble protein in the muscles of males and females. Patterns of changes in the amount of muscle water-soluble proteins and total lipids were recorded in the entire sample of the Caspian atherina during the pre-spawning period and at the beginning of spawning. This is typical for portioned spawning fish, in which the energy expenditure of the body is directed to the formation, maturation of gonads, the removal of sexual products and the maturation of the next portions during the spawning period. In September-October, that is, already at the stage of feeding, a multidirectional character in lipid and protein metabolism was noted in the body of atherina. According to the results of histological examination, the beginning of spawning in atherina was noted in April. Mature oocytes with resorption at an average level of 30-31% were recorded in the ovaries of portioned spawning atherina in February-April, which characterized the adaptive mechanisms of the organism in the conditions of this habitat. In September, there was a significantly positive (p < 0.05) statistical correlation between the concentration of total muscle lipids and the mass of fish (r = +0.47), which characterized a positive energy balance in the body, so necessary for the next stages of life.
atherina, muscles, gonads, total lipids, water-soluble protein
Introduction
The Caspian form of atherina (Atherina boyeri caspia Risso, 1810) is widespread throughout the Caspian Sea. It is a marine euryhaline fish species [1] that lives in desalinated pre-estuarine areas, in river mouths [2], as well as in its most saline areas. Recently, atherina in the Caspian Sea, in terms of its mass, occupies one of the second place after the Caspian sprat.
Atherina is the main object of by-catch in fishing Caspian sprat with fixed nets. In recent years, an increase in by-catch of atherina has been recorded in catches of Caspian sprat [3].
There is information in the literature about the size, weight and age characteristics of the spawning part of the atherina population in the modern ecological conditions of the Caspian Sea. The issues of atherina nutrition and variability of its morphometric features have been studied [4-6].
However, in the literature there are only fragmentary data on morphological and biochemical parameters of Caspian atherina and there is no data on its physiological capabilities, especially on the metabolic processes of the body that change during the period of individual development.
The purpose of the work is to undertake physiological and biochemical studies of atherina at various stages of the annual cycle in the Caspian Sea.
Research materials and methods
The work was carried out in the West Caspian Department of the Volga-Caspian Branch of the Federal State Budgetary Institution “VNIRO” (CaspNIRKH) in the period of 2014-2021. The research was carried out in the waters of the Kizlyar Bay and the adjacent area between the Bryansk and Suyutkino spit, on the Krainovsky coast from the Suyutkino spit in the south to the northern tip of Chechen Island.
Sampling was carried out in accordance with the instructions [7] in February, March, April, July, September, and October. The mass, length and gender were determined according to the generally accepted method [8]. The total lipid content in the muscles was determined by a modified method with phosphorus-vanillin reagent according to Zollner–Kirsch [9], the Warburg and Christian spectrophotometric method was used to determine the concentration of water-soluble protein [10, 11]. The determination of the stages of maturity of the gonads (NWG) of females was carried out by standard histological methods [12]. The results of the obtained data are presented in the form
of average values and their errors (M ± m). Statistical processing was carried out, according to the modern data processing rules [13, 14].
Research results and their discussion
The average linear-weight parameters of atherina (8.4 cm and 5.5 g) in 2021 were close to the average long-term indicators for the last 5 years (2016-2020). The average Fulton’s stomach fullness index was 0.944, which indicates a high feeding intensity of fish. During the study period, the predominance of females in the gender composition of the atherina population was noted – from 56 to 59% (Table 1).
Table 1
Biostatistical indexes of atherina in 2016-2021
|
Years |
Total catch, t |
Сatch on |
Average length, cm |
Average weight, g |
Proportion of females, % |
Fulton’s stomach |
|
2016 |
48.4 |
6.0 |
8.2 |
5.2 |
58 |
0.943 |
|
2017 |
34.6 |
3.5 |
8.3 |
5.4 |
57 |
0.944 |
|
2018 |
60.6 |
5.9 |
8.1 |
5.2 |
59 |
0.978 |
|
2019 |
57.6 |
5.3 |
8.4 |
5.5 |
55 |
0.928 |
|
2020 |
66.1 |
3.7 |
8.3 |
5.4 |
56 |
0.944 |
|
2021 |
70.7 |
2.7 |
8.4 |
5.5 |
58 |
0.927 |
|
Average |
56.3 |
4.5 |
8.3 |
5.4 |
57.2 |
0.944 |
In the course of the research the size and weight characteristics of atherina caught in different seasons of 2022 in the Middle Caspian region were determined (Table 2).
Table 2
Mass and length of the studied individuals of atherina in the Caspian Sea in 2022
|
Indexes |
Months |
|||||
|
February |
March |
April |
July |
September |
October |
|
|
Mass, g |
4.2 ± 0.2 |
4.2 ± 0.6 |
4.6 ± 0.4 |
6.5 ± 0.3 |
6.2 ± 0.2 |
6.5 ± 0.3 |
|
Length, cm |
8.2 ± 0.1 |
9.3 ± 0.4 |
9.8 ± 0.3 |
9.8 ± 0.2 |
10.1 ± 0.1 |
10.1 ± 0.2 |
In a living organism there take part constant metabolic processes characterized by the synthesis and decomposition of various biochemical compounds. To obtain information about synthesis and decomposition it is necessary to study the content of biochemical substrates of the organism aimed at the vital activity of cells and tissues in different periods of the annual cycle [15, 16].
Proteins and lipids are the main material substrate in animal tissues, the change of which characterizes plastic and energy metabolism [16].
Caspian atherina caught in February-March 2022 was characterized by a pre-spawning condition in April fish spawned.
In the pre-spawning period of February-March the results of studies of the amount of total lipids and water-soluble protein in the muscles of male and female fish did not reveal statistically significant differences. At the same time, the lipid content in females averaged 3.05%, in males – 3.08%, the concentration of water-soluble protein was 55.49 and 54.76 mg/g respectively (Table 3).
Table 3
Content of biochemical parameters in the muscles of atherina in the pre-spawning period
|
Gender |
Total lipids, % |
Water-soluble protein, mg/g |
|
Females |
3.05 ± 0.24 |
55.49 ± 4.42 |
|
Males |
3.08 ± 0.20 |
54.76 ± 2.16 |
In the absence of significant differences in the content of lipids and water-soluble protein in the muscles of females and males the analysis of physiological and biochemical parameters in individuals without sexual division is further presented in the work.
The lipid content in fish muscles is highly variable during the annual cycle [17-19]. The results of the study of total lipids and water-soluble protein in atherina muscle tissue from February to October are shown in Fig. 1.
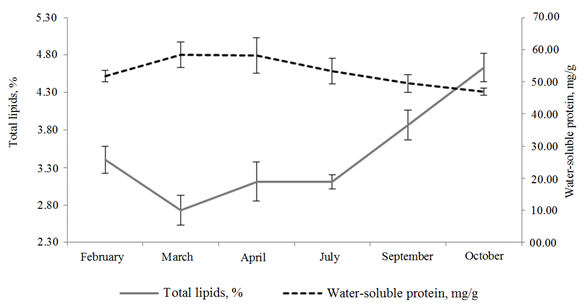
Fig. 1. Dynamics of the amount of muscle protein and total lipids
in the body of atherina from February to October 2022
In April there were noted fish that had already swept out the first portion of sexual products. The content of total lipids in the muscles of atherina was registered in the amount of 3.11%. Thus, in the period from March to April an increase in total lipids in the muscles was noted, since the main forming processes in the gonads had already ended, and during the transition of the gonads from stage IV to stage V of gonad maturation (SGM), mainly only their watering occurs [17]. During the spawning period the mobilization of lipids of atherina occurs, i. e. their accumulation. According to the literature data [4], the sexually mature atherina living in the Middle Caspian Sea does not have empty intestines in the spring and summer periods, i. e., a stable energy balance is established. Obviously, this is due to the process of shedding of roe and milt, which require significant energy consumption, as well as an increase in the mobility of fish [17]. In the Middle Caspian Sea atherina spawning begins in spring and lasts almost all summer [4]. In July the total lipid content in the muscles of atherina was at the level of April parameters – 3.11%.
In September-October the accumulation of lipid reserves occurs in the muscle tissue of atherina after spawning. During the feeding period a positive energy balance was registered, an increase in total lipids was noted to 3.87% in September and to 4.63% in October, which is associated with intensive food consumption. According to the literature data, large-size atherina in the Middle Caspian Sea do not have empty intestines in the spring-summer period, i.e. an increase in total muscle lipids in the autumn period is the result of fish nourishment earlier [4].
The statistical analysis revealed significant differences (p < 0.05) in the increase in the content of total lipids in the muscles of atherina in September, October in the feeding period relative to March by 29% and 41%, respectively, because in the spring period energy consumption for the formation and maturation of sexual products is carried out in the organism of atherina. In the autumn feeding period (October) there was a significant accumulation (p < 0.05) of total lipids compared to the pre-spawning and spawning stages (February-July), which is ensured by intensive food consumption.
With reference to total lipids, the amount of muscle water-soluble protein in atherina had a multidirectional character in different periods of the study.
In March in the pre-spawning period there was a slight increase in muscle protein to 58.40 mg/g compared to February (51.77 mg/g), which is explained by the predominance of protein synthesis over its decomposition. In April, with the beginning of spawning, the amount of water-soluble protein almost did not change and amounted to 58.17 mg/g. In July a decrease in protein in the muscle tissue of fish was revealed to 53.38 mg/g, due to its use as an energy substrate and removal from the body of atherina as part of sexual products. Similar metabolic mechanisms were noted in other fish with portioned spawning [17-19], which is explained by the redistribution of protein from the gonads to the muscles during the shedding of a repeated portion of the atherina roe. In September and October the amount of water-soluble protein does not increase, it slightly decreased to 49.51 and 46.94 mg/g, respectively, despite the consumption of food by atherina at these stages [4]. During the feeding period part of the protein in atherina is transformed into lipids, which accumulate intensively. During the studied periods of annual cycles there were no significant differences in the content of water-soluble protein in the muscle tissue of fish.
Thus, in the post-spawning – feeding period different directions of lipid and protein metabolism in the muscles were noted in atherina. Similar metabolic processes were registered at the feeding stage in Caspian sprat [18] and Azov anchovy [17].
In September a positive reliable (p < 0.05) statistical correlation was noted between the concentration of total muscle lipids and the mass of fish (r = +0.47), which characterized a positive energy balance in the organism necessary for the next stages of fish life.
Caspian atherina refers to fish species with portioned spawning, which causes a long duration of spawning and provides different possibilities to increase fertility. There exists some information in the literature about the fertility of atherina [20, 21], its high reliable correlation with the length and body weight of fish [4].
The reproduction of fish is based on the processes of development of the reproductive system – gametogenesis. The reproductive system of atherina in the Caspian Sea is currently poorly studied.
The results of the study of fragments of the gonads of female atherina in February-March are shown in Fig. 2.
|
|
|
|
a |
b |
|
|
|
|
c |
|
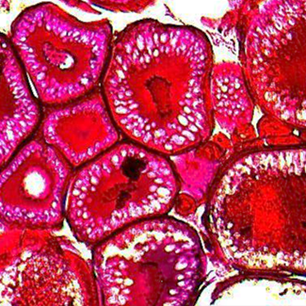
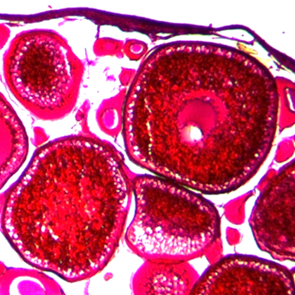
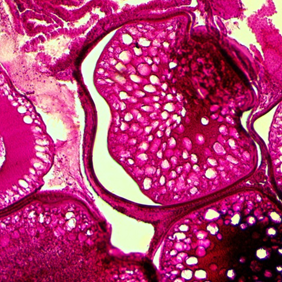
Fig. 2. Atherina oocytes at different stages of gonad maturation (SGM):
a – SGM III-IV; b – SGM IV; c – resorption at SGM IV. Magnification 10 × 4
In February fish with completed SGM IV predominated (46%). Individuals in the amount of 34% had SGM III-IV and 20% of fish had SGM III-IV. At the same time, resorbing oocytes of the period of trophoplasmic growth at an average level of 31% were noted in some species of atherina. The proportion of resorbing oocytes varied in the range of 17-47%.
In April the histological examination of fragments of the gonads of female atherina revealed that the largest number of fish (66%) were on SGM IV, individuals (20%) with gonads on SGM III-IV were noted. During this period atherina females (7%), that have started spawning, with gonads on SGM V, as well as individuals with ovaries on SGM VI-III (7%) were registered. Disorders of the development of maturing roe in the range from 14 to 44% of oocytes with an average level of 30% were determined.
The individual analysis of female ovaries did not reveal mass resorption of oocytes. Thus, scattered ovarian resorption was recorded in atherina [22, 23], which was an adaptive mechanism of the organism [24, 25] allowing the fish to tolerate unfavorable conditions without losing the ability to reproduce.
According to literature data [26], fish with portioned spawning, if roe of the first portion was covered by mass resorption, have spawning, but it occurs already due to the second portion of roe and the loss of progeny can be in the range of 50-60%. In addition, unlike simultaneously spawning fish species, resorption does not have an inhibitory effect on the process of vitellogenesis in fish with portion spawning, i. e. two processes can simultaneously take place in the ovaries: generation and degeneration.
Therefore, the identified resorption processes in atherina are not pathological due to the fact that there is no effect on the development of other oocytes [23].
Conclusion
Therefore, morpho-physiological and biochemical studies of Caspian atherina were carried out at various stages of the annual cycle in the Caspian Sea.
According to the results of histological analysis, the beginning of spawning in atherina was registered in April. Mature oocytes with resorption at an average level of 30-31% were observed in the ovaries of portioned spawning atherina in February-April, which was the adaptive mechanism of the organism in this habitat.
In the pre-spawning period and at the beginning of spawning, changes in the quantitative composition of lipids and proteins characteristic of portioned spawning fish were noted aimed at spending the organism's energy for the formation, maturation of gonads, shedding of sexual products and ripening of the next portions of roe. At the stage of feeding (September-October) the amount of muscle water-soluble proteins and total lipids had a multidirectional character in the organism of atherina.
1. Haldinova N. A. Materialy po razmnozheniyu i razvitiyu ryb v osolonennyh zalivah Severnogo Kaspiya // Tr. VNII mor. ryb. hoz-va i okeanografii. 1951. T. 18. S. 99-126.
2. Berg L. S. Ryby presnyh vod SSSR i sopredel'nyh stran. M.: Izd-vo AN SSSR, 1949. T. 3. S. 1001-1004.
3. Taibov P. S., Kanieva N. A., Barabanov V. V., Magomedova K. M. Promyslovo-biologicheskaya harakteristika obyknovennoy kil'ki Clupeonella cultriventris caspia i ateriny Ather inamochon caspia v Tersko-Kaspiyskom rybohozyaystvennom podrayone (Respublika Dagestan) v 2021 g. // Biologicheskoe raznoobrazie: izuchenie, sohranenie, vosstanovlenie, racional'noe ispol'zovanie: materialy III Mezhdunar. nauch.-prakt. konf. (Kerch', 13-18 sentyabrya 2022 g.). Simferopol': OOO «Izdatel'stvo Tipografiya «Ari-al», 2022. S. 238-243.
4. Dzhabrailov Yu. M., Ustarbekova D. A,, Mahmudova N. N. Osobennosti biologii ateriny Atherina boyeri caspia (Eichwald, 1838) v zapadnoy chasti Srednego Kaspiya // Bioraznoobrazie flory i fauny Dagestana: materialy region. nauch.-prakt. konf. (Mahachkala, 20 marta 2012 g.). Mahachkala: Izd-vo DGPU, 2012. S. 108-110.
5. Taibov P. S., Ashumova S. G., Dacieva M. M., Medzhidova A. K. Morfoekologicheskaya harakteristika ateriny Atherina boyeri Eichwald, 1838, v Tersko-Kaspiyskom rybohozyaystvennom podrayone // Kompleksnye issledovaniya biologicheskih resursov yuzhnyh morey i rek: materialy III Mezhdunar. nauch.-prakt. konf. molodyh uchenyh. Astrahan', 2012. S. 111-113.
6. Dzhabrailov Yu. M., Ustarbekova D. A., Zurhaeva U. D. Izmenchivost' morfologicheskih priznakov ateriny Atherina boyeri caspia (Eichwald, 1838) // Bioraznoob-razie flory i fauny Dagestana: materialy region. nauch.-prakt. konf. (Mahachkala, 20 marta 2012 g.). Mahachkala: Izd-vo DGPU, 2012. S. 106-108.
7. Instrukcii po sboru i pervichnoy obrabotke materialov vodnyh bioresursov Kaspiyskogo basseyna i sredy ih obitaniya / pod red. G. A. Sudakova. Astrahan': Izd-vo KaspNIRH, 2011. 193 s.
8. Pravdin I. F. Rukovodstvo po izucheniyu ryb. M.: Pisch. prom-st', 1966. 374 c.
9. Metody veterinarnoy klinicheskoy laborator-noy diagnostiki: sprav. / pod red. prof. I. P. Kondra-hina. M.: KolosS, 2004. 520 s.
10. Metody biologii razvitiya. Eksperimental'no-embriologicheskie, molekulyarno-biologicheskie i citologicheskie / pod red. T. A. Detlaf. M.: Nauka, 1974. 619 s.
11. Prakticheskaya himiya belka / pod red. A. Darbre. M.: Mir, 1989. 621 s.
12. Mikodina E. V., Sedova M. A., Chmilevskiy D. A. Gistologiya dlya ihtiologov. Opyt i sovety. M.: Izd-vo VNIRO, 2009. 112 s.
13. Bavrina A. P. Sovremennye pravila primeneniya parametricheskih i neparametricheskih kriteriev v statisticheskom analize mediko-biologicheskih dannyh // V pomosch' issledovatelyu. 2021. № 1 (66). S. 64-73.
14. Bavrina A. P., Borisov I. B. Sovremennye pravila primeneniya korrelyacionnogo analiza // V pomosch' issledovatelyu. 2021. № 3 (68). S. 70-79.
15. Kizevetter I. V. Biohimiya syr'ya vodnogo proishozhdeniya. M.: Pisch. prom-st', 1973. 424 s.
16. Malyarevskaya A. Ya. Obmen veschestv u ryb v usloviyah antropogennogo evtrofirovaniya vodoemov. Kiev: Naukova dumka, 1979. 256 s.
17. Shul'man G. E. Fiziologo-biohimicheskie osobennosti godovyh ciklov ryb: monogr. M.: Pisch. prom-st', 1972. 368 s.
18. Kozlova N. V., Latunov A. A., Makarova E. G., Mahlun A. V. Fiziologo-biohimicheskie izmeneniya kaspiyskoy obyknovennoy kil'ki (Clupeonella delicatula caspia Svetovidov) v razlichnye periody godovogo cikla // Vestn. Astrahan. gos. tehn. un-ta. Ser.: Rybnoe hozyaystvo. 2021. № 2. S. 116-124.
19. Belyanina T. N. Sezonnye izmeneniya zhirnosti belomorskoy koryushki v svyazi s sozrevaniem gonad // Zakonomernosti chislennosti ryb Belogo morya. M.: Nauka, 1966. S. 156-180.
20. Kazancheev E. N. Ryby Kaspiyskogo morya (opre-delitel'). M.: Leg. i pisch. prom-t', 1981. 167 s.
21. Yusufova Z. A. Znachenie ateriny Atherina mochon pontica natio caspia Eichwald v ispol'zovanii kormovoy bazy Kaspiyskogo morya: avtoref. dis. … kand. biol. nauk. Baku, 1969. 17 s.
22. Trusov V. E. Biologicheskaya harakteristika i puti rybohozyaystvennogo ispol'zovaniya osetrovyh, skaplivayuschihsya u Volgogradskoy plotiny // Osetrovye hozyaystva v vodoemah SSSR. M.: Izd-vo AN SSSR, 1997. S. 143-150.
23. Kurdyaeva V. P., Shapavalov M. E. O rezorbcii oocitov i narusheniyah v razvitii vosproizvoditel'noy sistemy u predstaviteley podsemeystva Sultrinae (semeystvo Ciprinidae) // Izv. Tihookean. nauch.-issledovat. rybohozyaystv. centra. 2002. T. 131. S. 390-408.
24. Gerbil'skiy N. L. Ekologo-gistologicheskoe napravlenie v ihtiologicheskih issledovaniyah // Tr. Vsesoyuz. sovesch. po biolog. osnovam rybovodstva. Tomsk, 1959. S. 31-36.
25. Faleeva T. I. Analiz atrezii oocitov u ryb v svyazi s adaptivnym znacheniem etogo yavleniya // Vopr. ihtiologii. 1965. T. 5. Vyp. 3 (36). S. 455-470.
26. Barhalov R. M. Osobennosti razvitiya i funkcionirovaniya reproduktivnoy sistemy ryb (na primere semeystva Cyprinidae) v izmenivshihsya ekologicheskih usloviyah vodoemov Severo-Zapadnogo Kaspiya: avtoref. dis. … kand. biol. nauk. Mahachkala, 2005. 26 s.
