











Russian Federation
Russian Federation
With habitats shrinking (e.g. due to construction projects that prevent access to areas upper the Volgograd Reservoir), changes in the environment, or illegal fishing in some areas, there is registered a sharp decrease of the population of Volga lamprey. Studying the histomorphological structure of this species has a significant impact on the prediction of changes in the population and on the assessment of the state of their internal organization. Problems in such sections as comparative embryology and histology of Volga lamprey have been poorly studied, which complicates the research. The aim of the research is to study the morphofunctional features of the organs of Caspian lamprey. The object of the study is Caspian (Volga) lamprey (Caspiomyzon wagneri), which is endemic to the Caspian Sea basin. The organs and tissues of lamprey were treated for the study in accordance with the methods generally accepted in histology. Microscopy of fixed, stained preparations was performed using a light microscope with immersion. Lampreys differ from other fish. The structure of the gill apparatus, the skin structural features, the structure of the digestive system organs, the structural features of the organs of the excretory and reproductive systems have been considered. The most intense pathological processes were characteristic of the gills, as the organ actively contacts with the environment. The studied species showed signs of epithelial hyperplasia, vascular disorders, and a change in the shape of lamellae.
Caspian lamprey, gills, kidneys, intestine, gonads, lamellae
Введение
В настоящее время влияние промысловой деятельности человека на рыбные запасы настолько велико, что не только их увеличение, но и поддержание стабильного уровня является важной задачей, решение которой требует научного подхода к разработке рекомендаций как по их использованию, так и по их охране. Каспийская минога (Caspiomyzon wagneri (Kessler)) – единственный вид из класса круглоротых в бассейне Каспийского моря, который был ценным объектом промысла до 1997 г.
В XIX в. и первой половине XX в. волжскую миногу добывали для использования в качестве освещения (сушеных миног сжигали вместо свечей). В 60-х гг. ее использовали для вытопки жира. После того каспийскую миногу стали употреблять в пищу. У миноги отсутствуют желчь с горьким вкусом, остатки пищи в кишечнике и костный скелет, что позволяет употреблять каспийскую миногу целиком без отходов [1].
По пищевой ценности миноге почти нет равных в Волжско-Каспийском бассейне. Свежая минога содержит около 29 % жира, 12 % белковых веществ, 2 % минеральных солей, 58 % влаги. Жир миноги содержит много йода, полный набор незаменимых аминокислот, витамины А, В, В12, С, D1, Е и др., он очень полезен и приятен на вкус.
Наблюдавшиеся в последние годы регулирование Волги и интенсивные зимние пропуски воды с Волжской гидроэлектростанции, влияющие на скорость течения, состояние нерестилищ миноги и ход ее нереста, а также возможный незаконный промысел миноги в некоторых районах сильно повлияли на сокращение ее численности. Изучение состояния гистоморфологической структуры органов данного вида оказывает значительное влияние на прогнозирование изменения численности популяции. Вопросы в таких разделах, как сравнительная эмбриология, гистология волжской миноги, изучены слабо, что затрудняет исследования [2].
Объект исследования – каспийская (волжская) минога (Caspiomyzon wagneri), являющаяся эндемиком бассейна Каспийского моря с проходным образом жизни и относящаяся к группе водных хордовых [3, 4].
Цель работы – изучение морфофункциональных особенностей органов каспийской миноги.
Материалы и методы
Для гистологического анализа органы и ткани волжской миноги (Caspiomyzon wagneri) фиксировали в 10 %-м нейтральном формалине. Материал обработан в соответствии с общепринятыми методами [5, 6]. Пробы органов заливали парафином, были сделаны срезы толщиной 4–5 мкм, которые затем окрашивали гематоксилин-эозином. Всего было изготовлено и проанализировано 180 срезов тканей и органов.
Микроскопию фиксированных окрашенных препаратов проводили при помощи светового микроскопа с применением иммерсии. Микрофотосъемка срезов органов произведена с помощью фотонасадки SONI DSC-W7 [7].
Особенности строения жаберного аппарата каспийской миноги
В ходе исследований было выявлено, что взрослые миноги имеют по 7 расположенных по бокам жаберных отверстий, которые ведут к жаберным мешочкам. Внешние отверстия поддерживаются хрящом, который расположен под кожным покровом, т. к. жаберных дуг нет [2, 8, 9]. Отмечено плотное расположение жаберных филаментов с тонкими удлиненными ламеллами во внутренней стороне жаберных мешков (рис. 1).
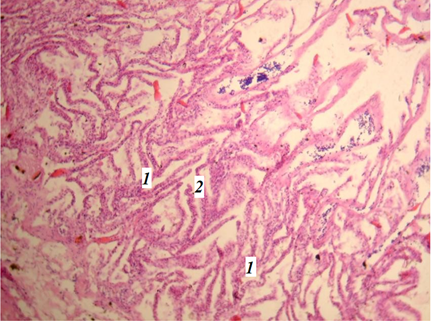
Рис. 1. Фрагмент жабр миноги (гематоксилин-эозин; ув. 400):
1 – филаменты жабр; 2 – ламеллы жабр
Fig. 1. Fragment of the lamprey gills (hematoxylin-eosin; magnification 400):
1 – filaments of gills; 2 – lamellae of gills
Стенки жаберных мешков содержат только соединительную ткань, кольцевой слой поперечнополосатой мускулатуры расположен вокруг жаберных отверстий. Жаберные мешки эндотермического происхождения, с многочисленными жаберными лепестками, расположенными во внутренних стенках.
Было выявлено, что ламеллы у каспийской миноги сильно вытянутые и извитые. Более того, длина ламелл основания и вершины филамента короче центральных. Сами ламеллы неправильной формы: изогнутые, неравномерной толщины, с расширенными дистальными участками.
Часто была обнаружена гиперплазия эпителия дыхательных путей (рис. 2).
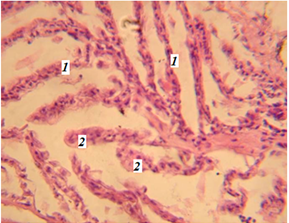
Рис. 2. Фрагмент жабр миноги (гематоксилин-эозин; ув. 800):
1 – ламеллы жабр; 2 – гиперплазия респираторного эпителия ламелл
Fig. 2. Fragment of the lamprey gills (hematoxylin-eosin; magn. 800):
1 – lamellae of gills; 2 – hyperplasia of the respiratory epithelium of lamellae
Концевые участки ламелл имели булавовидные расширения. Пронизывающие их сосуды, расположенные в терминальной зоне, были значительно расширены и заполнены форменными элементами крови. Все вышеперечисленные изменения в организации жаберных ламелл – показатель морфофункциональной адаптации данного органа.
Особенности строения кожного покрова
Кожа миног гладкая, обогащенная одноклеточными железами, которые находятся в эпидермисе. По данной причине кожа миноги обильно покрыта слизью. Кожа построена из слоев эпителия и соединительной ткани [2].
Эпидермис представлен многослойным ороговевающим эпителием. В дерме есть 2 слоя: сосочковый находится непосредственно под эпидермисом, а сетчатый слой находится глубже. В сетчатом слое дермы большую часть пространства занимают толстые пучки коллагеновых волокон, что делает ткань плотной, бесформенной вследствие их беспорядочного направления. Также были зарегистрированы пигментсодержащие клетки – хроматофоры (рис. 3).

Рис. 3. Фрагмент кожи миноги (гематоксилин-эозин; ув. 400):
1 – эпидермис; 2 – коллагеновые волокна; 3 – хроматофоры
Fig. 3. Fragment of the lamprey skin(hematoxylin-eosin; magn. 400):
1 – epidermis; 2 – collagen fibers; 3 – chromatophores
Особенности структуры органов пищеварительной системы
Желудок у миног отсутствует, поэтому пищевод, огибающий сердце, переходит непосредственно в кишечник, отделенный клапаном [10]. Границы клеток были видны неточно. Стенки пищевода имеют маломощный мышечный слой. Ворсинки небольшой высоты с тонким слоем призматических эпителиальных клеток, слизистая оболочка образована рыхлой волокнистой соединительной тканью с редко встречающимися отдельными клетками и кровеносными капиллярами. В кишке также были зафиксированы отдельные гладкомышечные клетки, поднимающиеся в строму ворсинки. Слой мышечных клеток, образующих мышечную мембрану, был очень тонким [11, 12].
В полости кишечника обнаружена складка слизистой оболочки – спиральный клапан с наружной поверхностью из слоя эпителиальных клеток призматической формы, где были зафиксированы довольно крупные скопления гемопоэтической ткани, которые характеризовались наличием клеток крови всех рядов, с преобладанием лимфоидных клеток (рис. 4).
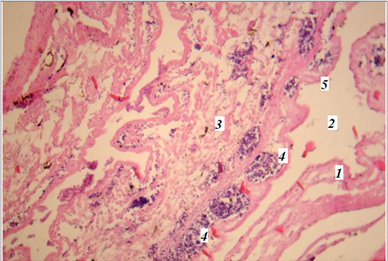
Рис. 4. Фрагмент пищеварительной системы миноги
(гематоксилин-эозин; ув. 400): 1 – стенка пищеварительной трубки;
2 – полость пищеварительной трубки; 3 – спиральный клапан; 4 – гемопоэтические образования; 5 – кишечный эпителий
Fig. 4. Fragment of the lamprey digestive system (hematoxylin-eosin: magn. 400):
1 – wall of the digestive tube; 2 – cavity of the digestive tube; 3 – spiral valve; 4 – hematopoietic formations; 5 – intestinal epithelium
Особенности строения органов выделительной системы
Первичные (туловищные) почки в форме лент сохраняются в качестве органов выделительной системы миног, расположены по бокам вдоль всей брюшной полости, начиная от спинной аорты. Почки так же, как и другие полостные органы, подвешены складкой к стенке, они наиболее мaссивны в заднем отделе, где отчетливо видны мочеточники – вольфовы каналы [13].
Вдоль почки, по нижнему ее краю, проходит мочеточник, который открывается самостоятельными отверстиями в мочеполовой синус. В почках были обнаружены почечные (мальпигиевы) тельца (округлые образования с высокой концентрацией клеток), что является явным признаком наличия коркового вещества. В корковом веществе также имеются почечные канальцы (рис. 5).
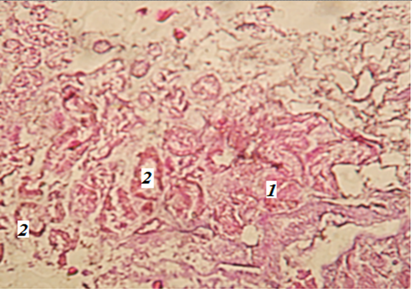
Рис. 5. Фрагмент почки миноги
(гематоксилин-эозин; ув. 400): 1 – почечное тельце; 2 – почечные канальцы
Fig. 5. Fragment of the lamprey kidney
(hematoxylin-eosin; magn. 400): 1 – renal body; 2 – renal tubules
Особенности строения половой системы
У круглоротых половые железы развиваются за счет генитальных складок, тянущихся вдоль полости тела. Они висят на специальных складках брюшины. У миноги половые железы непарные. Гонады в виде длинных лент висят над кишечником, заполняя почти всю полость тела. У них нет собственного выводного протока, и половые продукты попадают в полость тела, откуда выводятся наружу через генитальные поры за задним проходом. Лопастной семенник снаружи покрыт серозной прочной оболочкой. Полость органа заполнена многочисленными замкнутыми фолликулами, ограниченными соединительнотканной прослойкой (рис. 6).
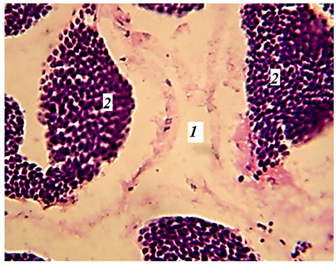
Рис. 6. Фрагмент гонад миноги (гематоксилин-эозин; ув. 400):
1 – извитые семенные канальцы; 2 – сперматиды
Fig. 6. Fragment of the lamprey gonads (hematoxylin-eosin; magn. 400):
1 – convoluted seminal tubules; 2 – spermatids
Семенные канальцы извивались в разных плоскостях без строгого порядка. В разрезе они представляли собой отдельные участки неправильной формы. Полости канальцев были заполнены крупными клетками – первоначальными семенными клетками – сперматогониями, которые интенсивно размножались.
Заключение
В процессе исследования гистоструктуры внутренних органов волжской миноги было установлено, что их строение имеет свои особенности. Выявлены патологические изменения органов миноги. Наиболее интенсивные патологические процессы были зарегистрированы для жабр как органа, наиболее активно контактирующего с окружающей средой. У исследуемых особей наблюдались признаки эпителиальной гиперплазии, сосудистые нарушения, изменение формы ламелл, что может быть вызвано неблагоприятными факторами среды обитания исследуемого объекта.
1. Ginzburg Ya. I. Nerestovaya populyaciya kaspiyskoy minogi posle zaregulirovaniya r. Volgi plotinoy Volgogradskoy GES // Vopr. ihtiologii. 1969. T. 9, vyp. 6/59. S. 1022-1031.
2. Suvorov E. K. Osnovy ihtiologii. L.: Sovet. nauka, 1948. 580 s.
3. Berg L. S. Ryby presnyh vod SSSR i sopredel'nyh stran. M.; L.: Izd-vo AN SSSR, 1948. T. 1. 468 s.
4. Holcík J. Caspiomyzon wagneri (Kessler, 1870) // The Freshwater fishes of Europe. 1986. V. 1, part I. P. 119-142.
5. Nazarov D. Yu., Kucheryavyy A. V., Pavlov D. S. Distribution and habitat types of the lamprey larvae in rivers across Eurasia // Jawless Fishes of the World. Cambridge: Cambridge Scholars Publ., 2016. V. 1. P. 280-298.
6. Shashulovskiy V. A., Ermolin V. P. Sostav ihtiofauny Volgogradskogo vodohranilischa // Vopr. ihtiologii. 2005. T. 45, № 3. S. 324-330.
7. Volkova O. V., Eleckiy Yu. K. Osnovy gistologii s gistologicheskoy tehnikoy. M.: Medicina, 1982. 304 s.
8. Matey V. E. Funkcional'naya morfologiya zhabernogo epiteliya presnovodnyh kostistyh ryb // Fiziologiya, biohimiya i toksikologiya presnovodnyh zhivotnyh. L.: Nauka, 1990. C. 104-141.
9. Pravdin I. F. Rukovodstvo po izucheniyu ryb. M.: Pisch. prom-st', 1966. 376 s.
10. Kottelat M., Freyhof J. Handbook of European freshwater fishes // Publications Kottelat, Cornol and Freyhof. Berlin, 2007. 646 p.
11. Fedorova N. N., Grushko M. P., Kanieva N. A. Patomorfologicheskie izmeneniya zhiznenno vazhnyh organov volzhskih ryb // Vestn. Astrahan. gos. tehn. un-ta. Ser.: Rybnoe hozyaystvo. 2019. № 4. C. 104-108.
12. Renaud C. B. Lampreys of the world. An annotated and illustrated catalogue of lamprey species known to date // FAO Species Catalogue for Fishery Purposes. Rome: FAO, 2011. N. 5. 109 p.
13. Richardson M. K., Admiraal J., Wright G. M. Developmental anatomy of lampreys // Biological reviews. 2010. V. 85. Iss. 1. P. 1-33.
